Guerriero Christopher J, Reutter Karl-Richard, Augustine Andrew A, Preston G Michael, Weiberth Kurt F, Mackie Timothy D, Cleveland-Rubeor Hillary C, Bethel Neville P, Callenberg Keith M, Nakatsukasa Kunio, Grabe Michael, Brodsky Jeffrey L
Department of Biological Sciences, University of Pittsburgh, Pittsburgh, PA 15260.
Cardiovascular Research Institute, Department of Pharmaceutical Chemistry, University of California, San Francisco, San Francisco, CA 94158.
Mol Biol Cell. 2017 Jul 15;28(15):2076-2090. doi: 10.1091/mbc.E17-03-0184. Epub 2017 May 24.
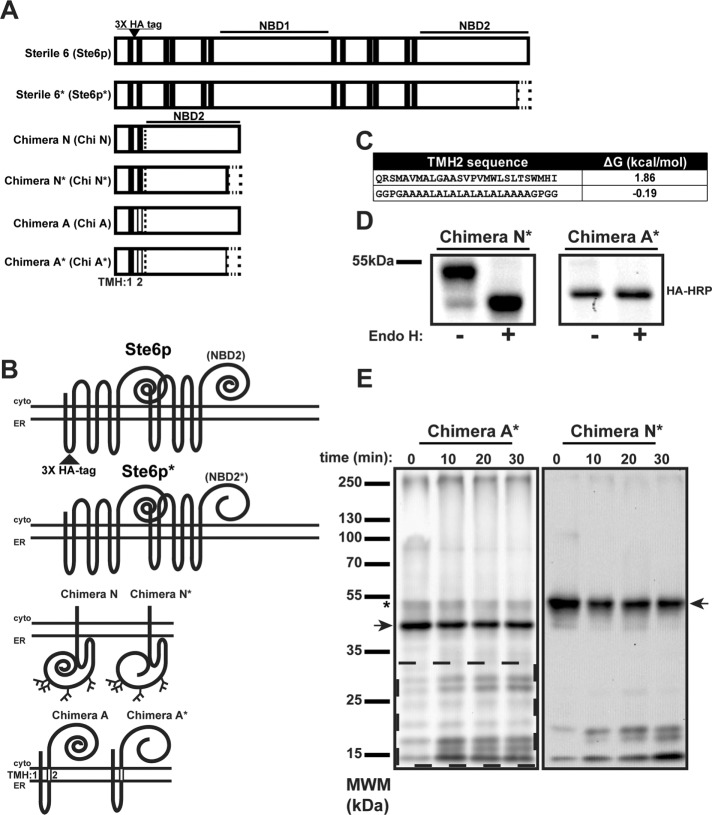
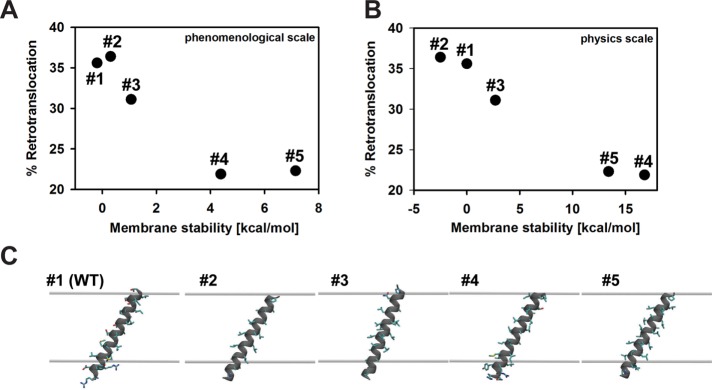
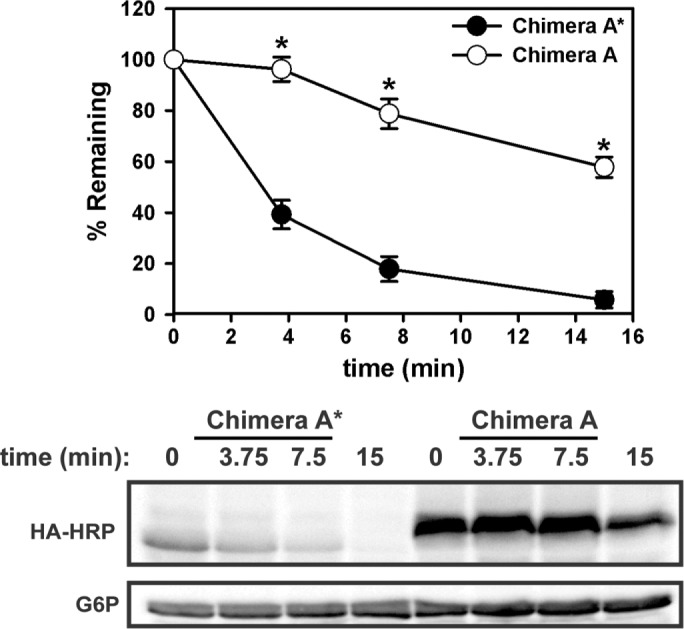
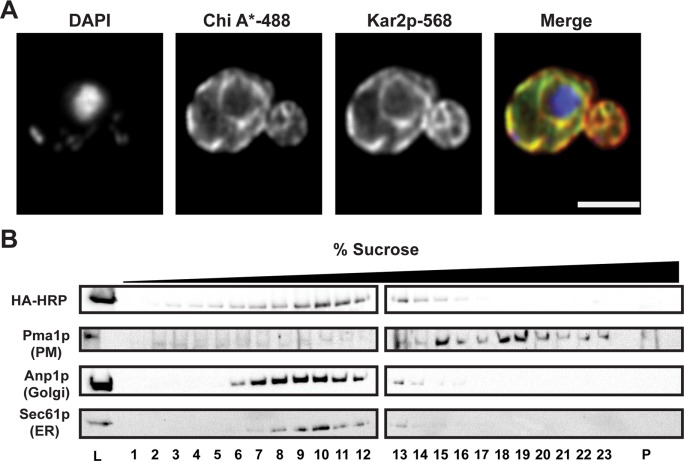
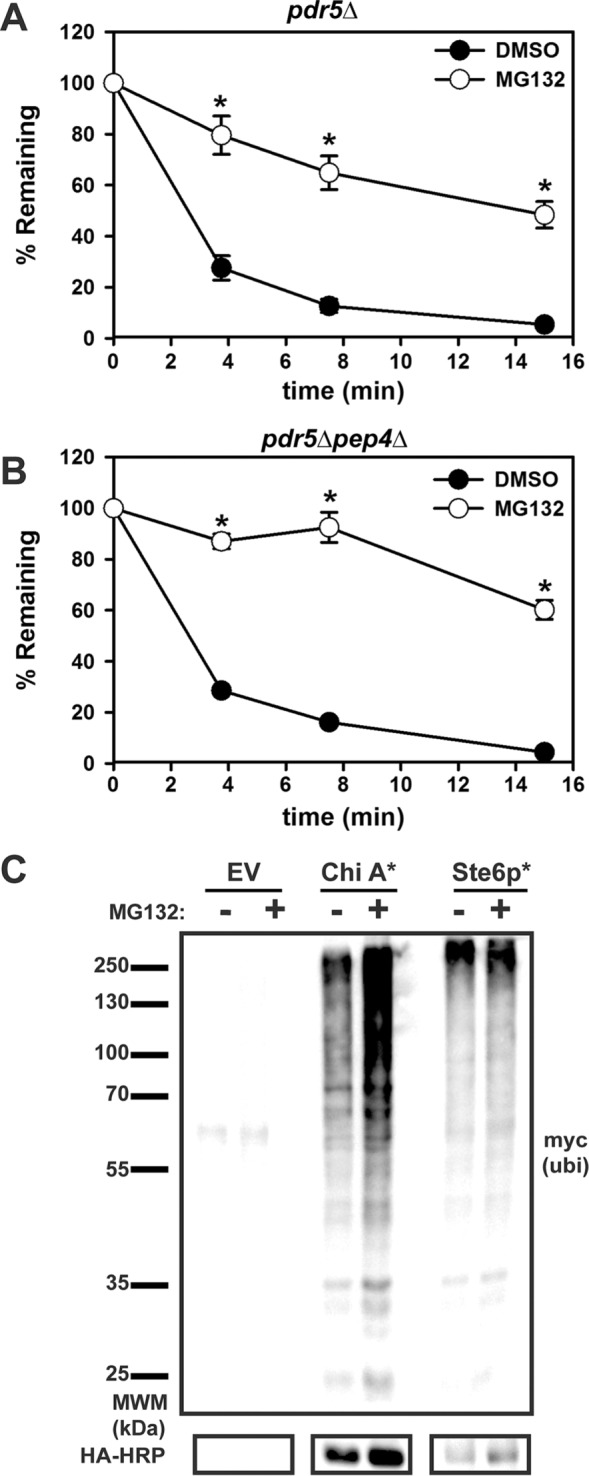
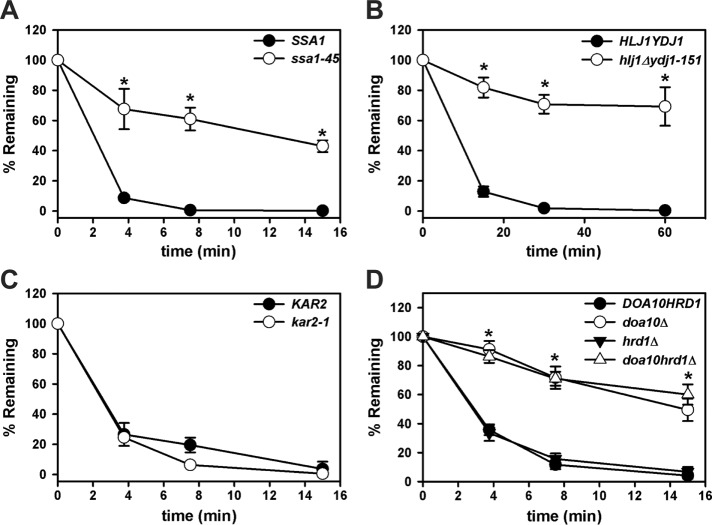
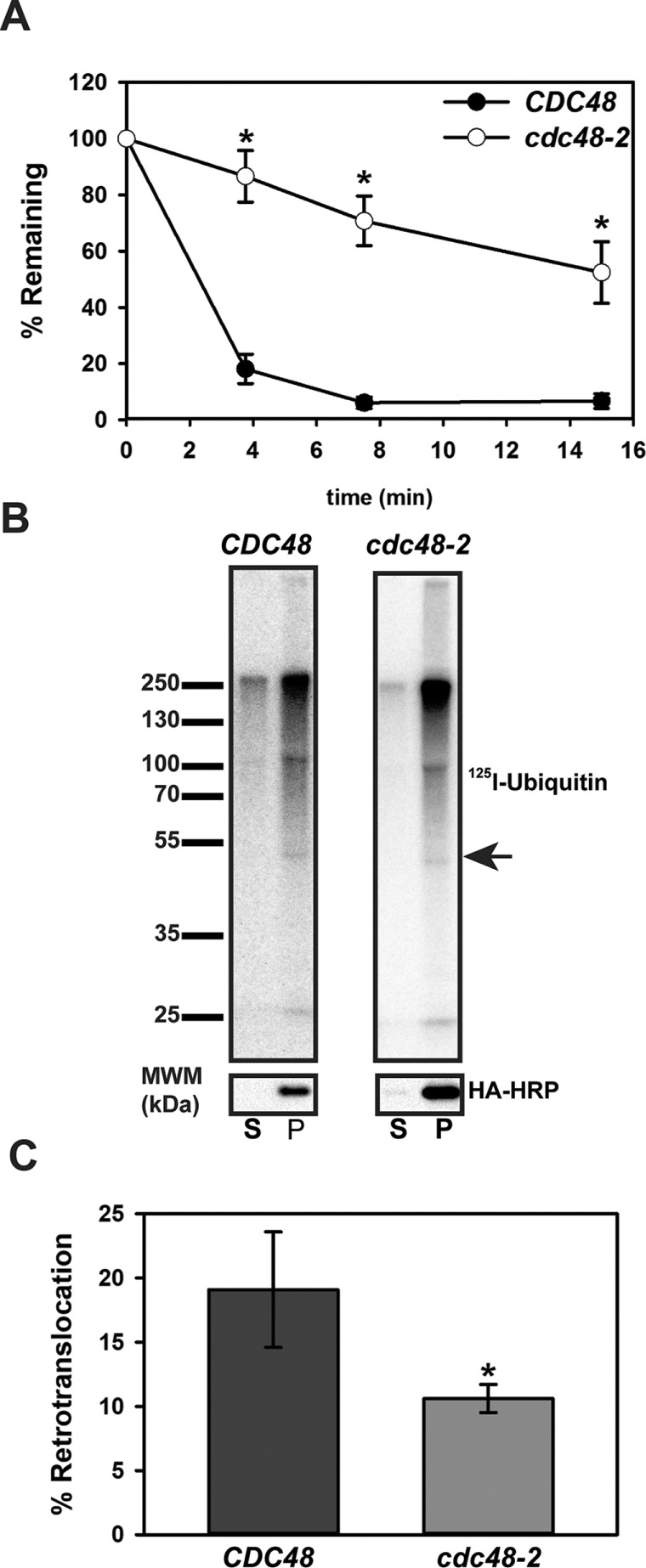
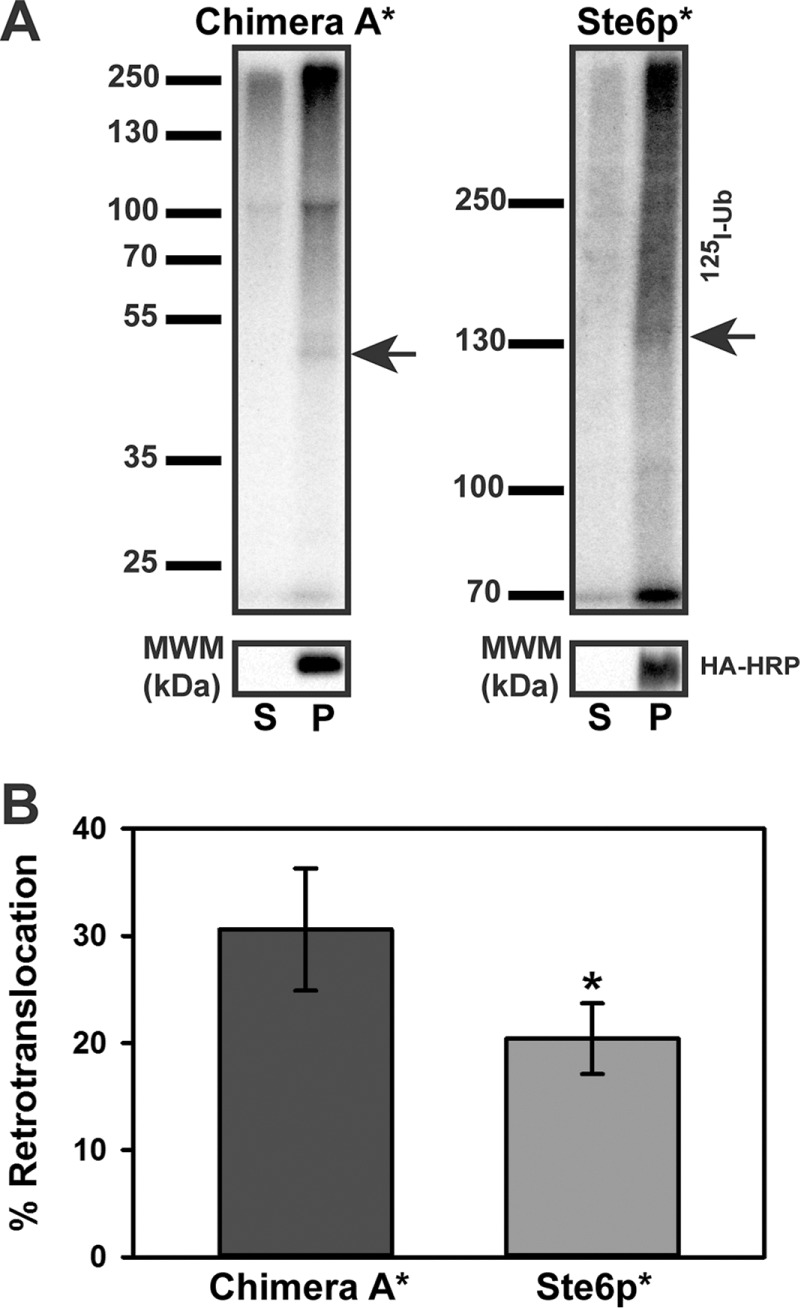
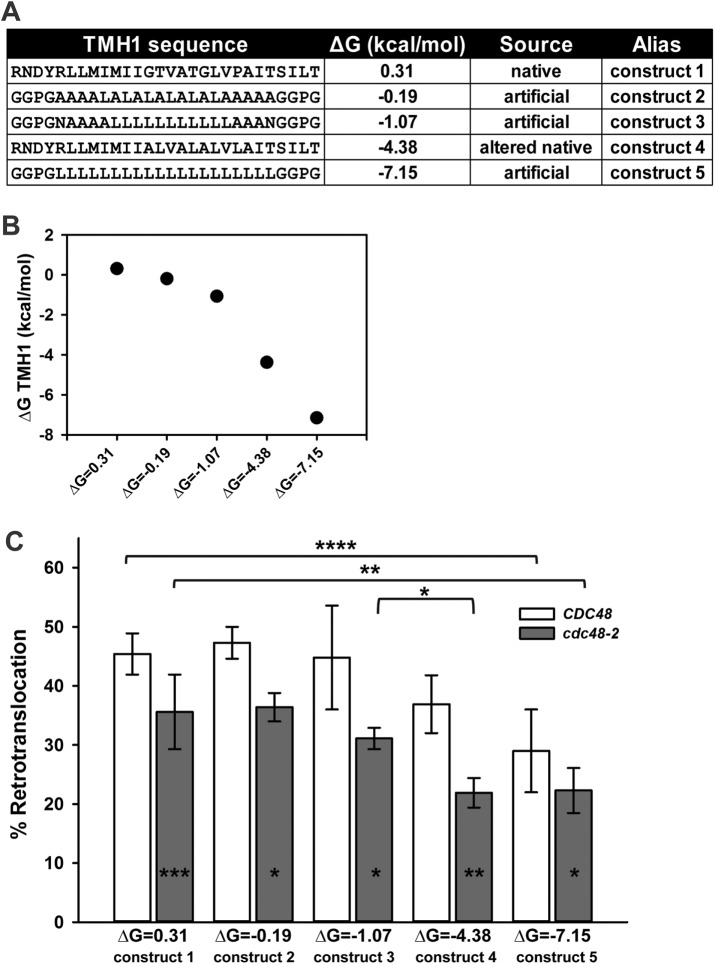
Integral membrane proteins fold inefficiently and are susceptible to turnover via the endoplasmic reticulum-associated degradation (ERAD) pathway. During ERAD, misfolded proteins are recognized by molecular chaperones, polyubiquitinated, and retrotranslocated to the cytoplasm for proteasomal degradation. Although many aspects of this pathway are defined, how transmembrane helices (TMHs) are removed from the membrane and into the cytoplasm before degradation is poorly understood. In this study, we asked whether the hydrophobic character of a TMH acts as an energetic barrier to retrotranslocation. To this end, we designed a dual-pass model ERAD substrate, Chimera A*, which contains the cytoplasmic misfolded domain from a characterized ERAD substrate, Sterile 6* (Ste6p*). We found that the degradation requirements for Chimera A* and Ste6p* are similar, but Chimera A* was retrotranslocated more efficiently than Ste6p* in an in vitro assay in which retrotranslocation can be quantified. We then constructed a series of Chimera A* variants containing synthetic TMHs with a range of Δ values for membrane insertion. TMH hydrophobicity correlated inversely with retrotranslocation efficiency, and in all cases, retrotranslocation remained Cdc48p dependent. These findings provide insight into the energetic restrictions on the retrotranslocation reaction, as well as a new computational approach to predict retrotranslocation efficiency.
整合膜蛋白折叠效率低下,且易通过内质网相关降解(ERAD)途径进行周转。在ERAD过程中,错误折叠的蛋白被分子伴侣识别、多聚泛素化,然后逆向转运到细胞质中进行蛋白酶体降解。尽管该途径的许多方面已被明确,但在降解之前跨膜螺旋(TMH)如何从膜上移除并进入细胞质却知之甚少。在本研究中,我们探究了TMH的疏水性是否构成逆向转运的能量障碍。为此,我们设计了一种双程模型ERAD底物Chimera A*,它包含来自已鉴定的ERAD底物不育6*(Ste6p*)的细胞质错误折叠结构域。我们发现Chimera A和Ste6p的降解要求相似,但在可定量逆向转运的体外试验中,Chimera A比Ste6p更有效地进行了逆向转运。然后,我们构建了一系列含有具有一系列膜插入Δ值的合成TMH的Chimera A*变体。TMH疏水性与逆向转运效率呈负相关,并且在所有情况下,逆向转运均依赖于Cdc48p。这些发现为逆向转运反应的能量限制提供了见解,以及一种预测逆向转运效率的新计算方法。