Department of Biochemistry, University of Toronto, Toronto, Ontario, Canada.
PLoS One. 2013;8(4):e60445. doi: 10.1371/journal.pone.0060445. Epub 2013 Apr 3.
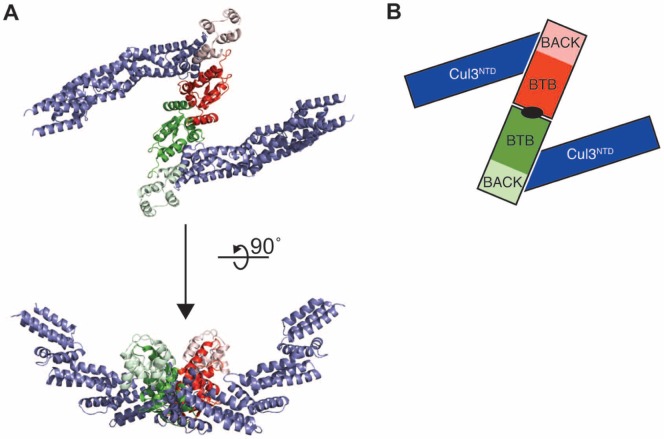
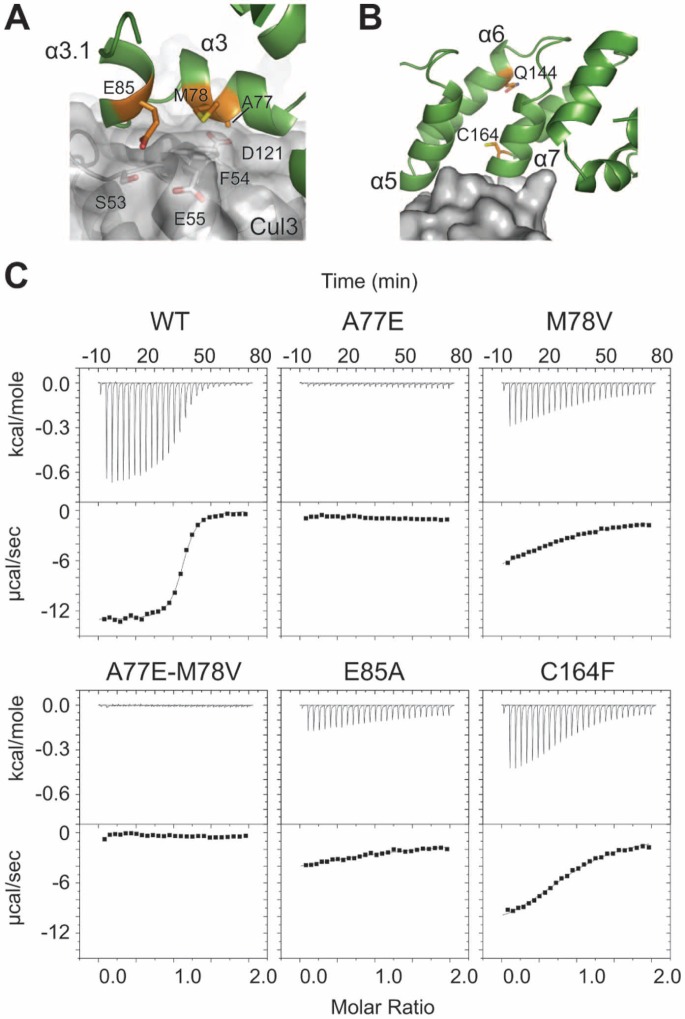
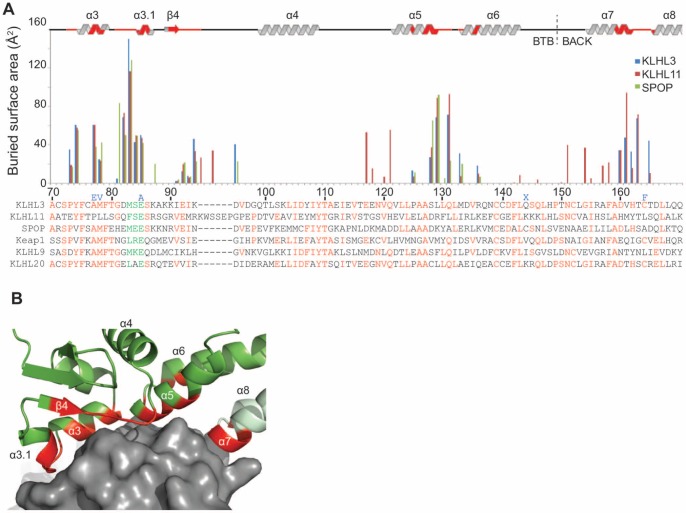
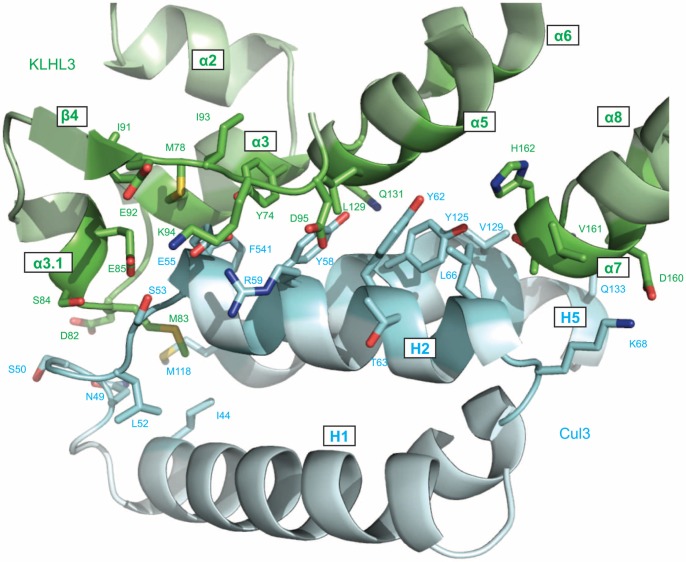
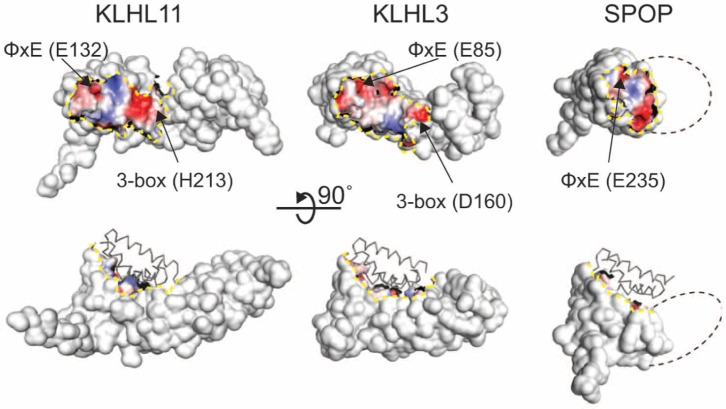
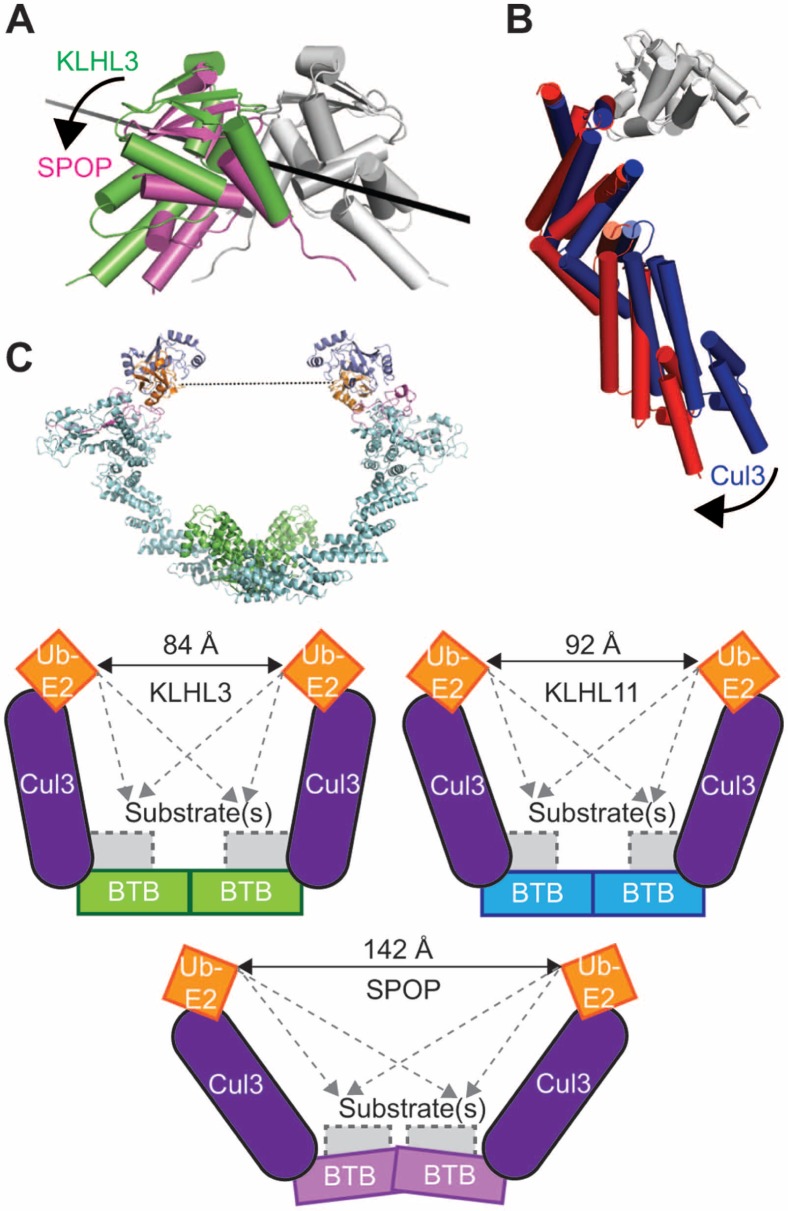
KLHL3 is a BTB-BACK-Kelch family protein that serves as a substrate adapter in Cullin3 (Cul3) E3 ubiquitin ligase complexes. KLHL3 is highly expressed in distal nephron tubules where it is involved in the regulation of electrolyte homeostasis and blood pressure. Mutations in KLHL3 have been identified in patients with inherited hypertension disorders, and several of the disease-associated mutations are located in the presumed Cul3 binding region. Here, we report the crystal structure of a complex between the KLHL3 BTB-BACK domain dimer and two copies of an N terminal fragment of Cul3. We use isothermal titration calorimetry to directly demonstrate that several of the disease mutations in the KLHL3 BTB-BACK domains disrupt the association with Cul3. Both the BTB and BACK domains contribute to the Cul3 interaction surface, and an extended model of the dimeric CRL3 complex places the two E2 binding sites in a suprafacial arrangement with respect to the presumed substrate-binding sites.
KLHL3 是一个 BTB-BACK-Kelch 家族蛋白,作为 Cullin3(Cul3)E3 泛素连接酶复合物中的底物衔接子。KLHL3 在远曲小管中高度表达,在那里它参与电解质稳态和血压的调节。KLHL3 的突变已在遗传性高血压疾病患者中被鉴定出来,并且几种与疾病相关的突变位于假定的 Cul3 结合区域。在这里,我们报告了 KLHL3 BTB-BACK 结构域二聚体和 Cul3 的两个 N 端片段之间复合物的晶体结构。我们使用等温滴定量热法直接证明 KLHL3 BTB-BACK 结构域中的几种疾病突变破坏了与 Cul3 的结合。BTB 和 BACK 结构域都有助于 Cul3 的相互作用表面,并且二聚体 CRL3 复合物的扩展模型将两个 E2 结合位点置于与假定的底物结合位点相对的表面排列。