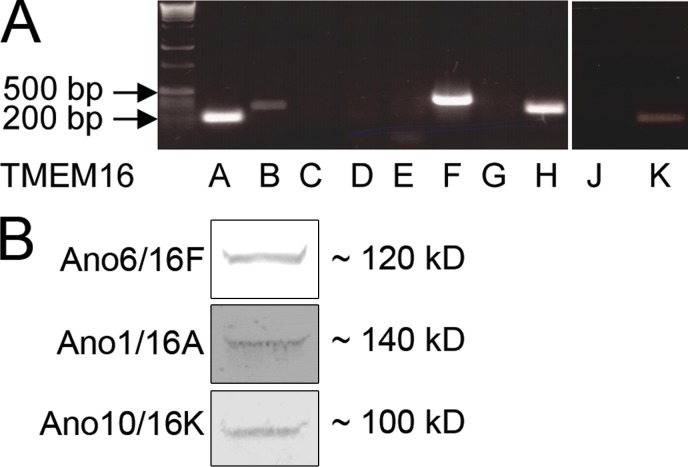
Department of Biology, University of Copenhagen, DK-2100 Copenhagen Ø, Denmark.
J Gen Physiol. 2013 May;141(5):585-600. doi: 10.1085/jgp.201210861.
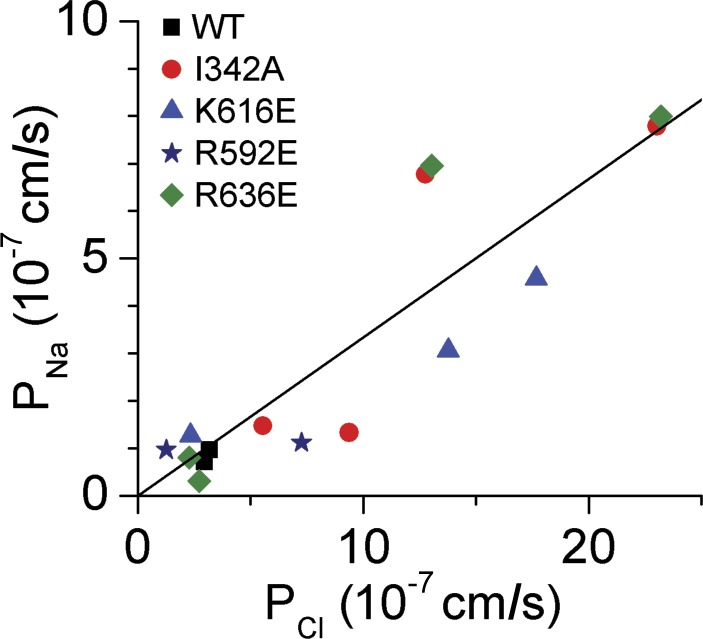
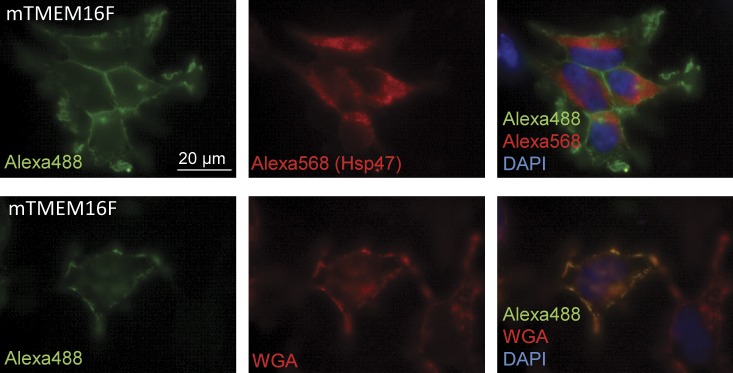
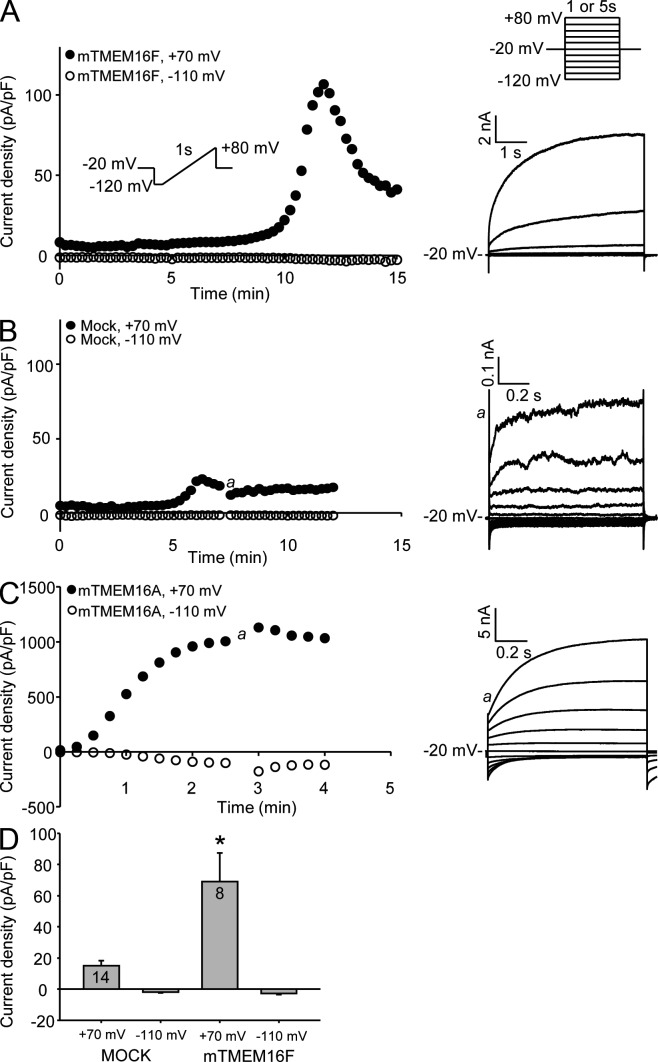
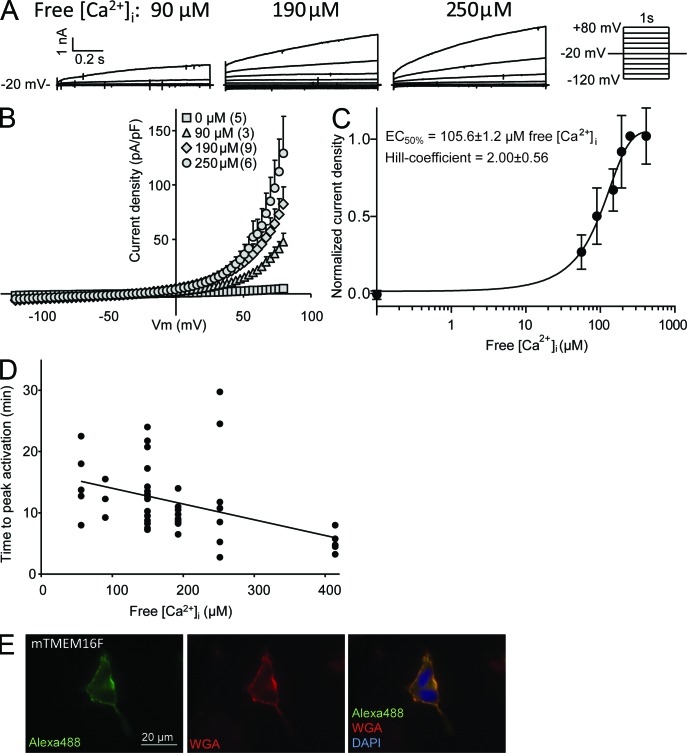
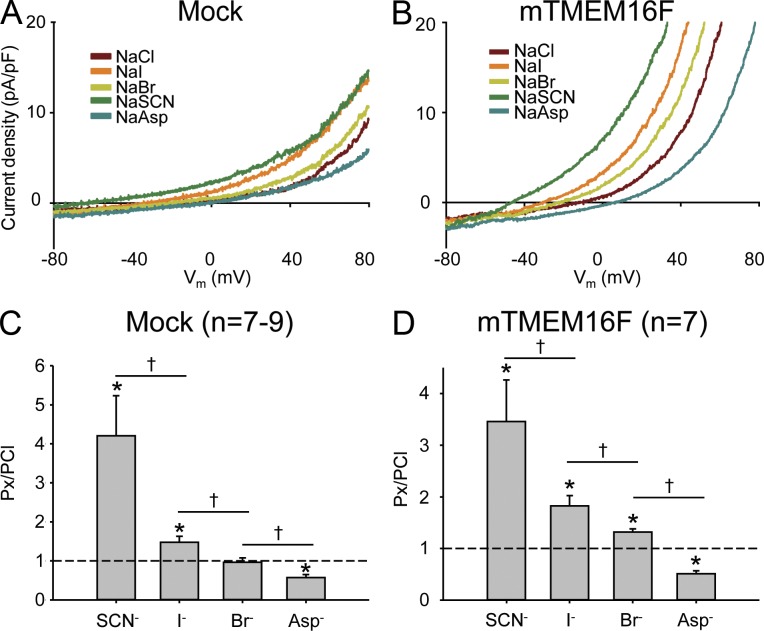
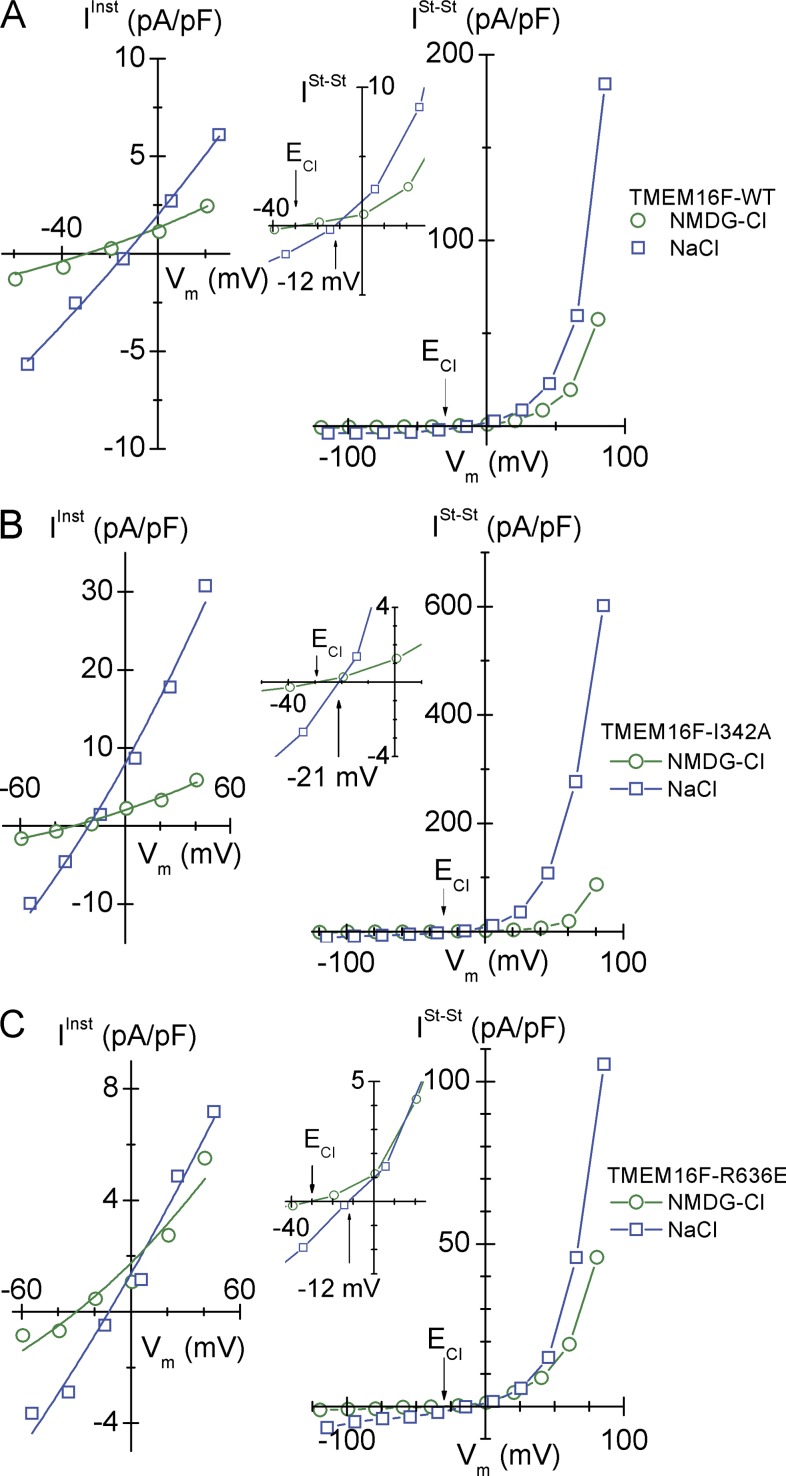
Members of the TMEM16 (Anoctamin) family of membrane proteins have been shown to be essential constituents of the Ca(2+)-activated Cl(-) channel (CaCC) in many cell types. In this study, we have investigated the electrophysiological properties of mouse TMEM16F. Heterologous expression of TMEM16F in HEK293 cells resulted in plasma membrane localization and an outwardly rectifying ICl,Ca that was activated with a delay of several minutes. Furthermore, a significant Na(+) current was activated, and the two permeabilities were correlated according to PNa = 0.3 PCl. The current showed an EC50 of 100 µM intracellular free Ca(2+) concentration and an Eisenman type 1 anion selectivity sequence of PSCN > PI > PBr > PCl > PAsp. The mTMEM16F-associated ICl,Ca was abolished in one mutant of the putative pore region (R592E) but retained in two other mutants (K616E and R636E). The mutant K616E had a lower relative permeability to iodide, and the mutant R636E had an altered anion selectivity sequence (PSCN = PI = PBr = PCl > PAsp). Our data provide evidence that TMEM16F constitutes a Ca(2+)-activated anion channel or a pore-forming subunit of an anion channel with properties distinct from TMEM16A.
TMEM16(Anoctamin)家族的膜蛋白成员已被证明是许多细胞类型中 Ca2+激活的 Cl-通道(CaCC)的基本组成部分。在这项研究中,我们研究了小鼠 TMEM16F 的电生理特性。TMEM16F 在 HEK293 细胞中的异表达导致质膜定位和外向整流的 ICl,Ca,其延迟几分钟后被激活。此外,还激活了显著的 Na+电流,并且两种通透性根据 PNa = 0.3 PCl 相关。电流的 EC50 为 100 µM 细胞内游离 Ca2+浓度,Eisenman 类型 1 阴离子选择性序列为 PSCN > PI > PBr > PCl > PAsp。在假定的孔区(R592E)的一个突变体中,mTMEM16F 相关的 ICl,Ca 被消除,但在另外两个突变体(K616E 和 R636E)中保留。突变体 K616E 对碘化物的相对通透性较低,而突变体 R636E 的阴离子选择性序列发生改变(PSCN = PI = PBr = PCl > PAsp)。我们的数据提供了证据,表明 TMEM16F 构成了 Ca2+激活的阴离子通道或阴离子通道的孔形成亚基,其特性与 TMEM16A 不同。