Graduate Group of Pharmacology and Toxicology, University of California, Davis, Davis, CA.
Center for Neuroscience, University of California, Davis, Davis, CA.
J Gen Physiol. 2019 Apr 1;151(4):518-531. doi: 10.1085/jgp.201812270. Epub 2019 Jan 22.
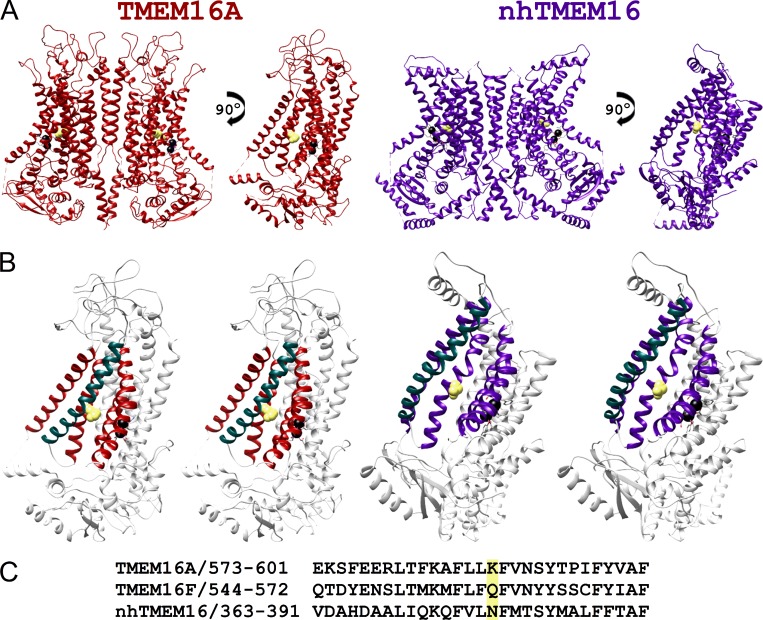
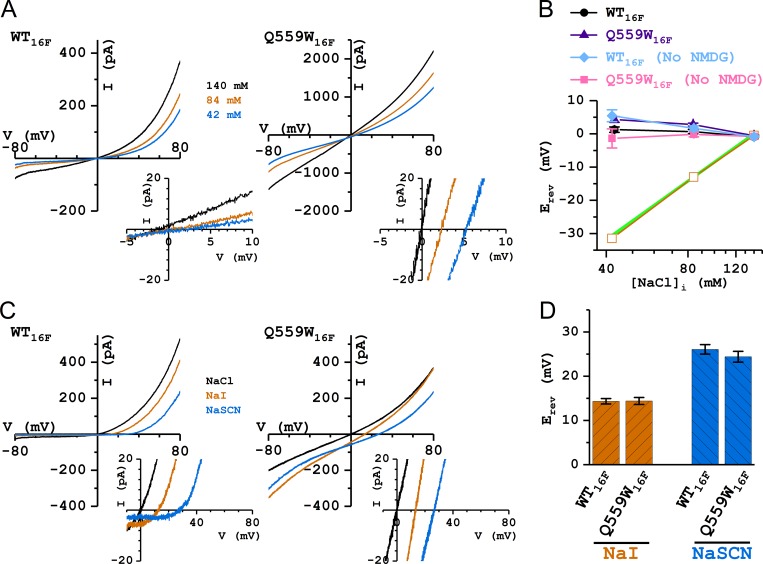
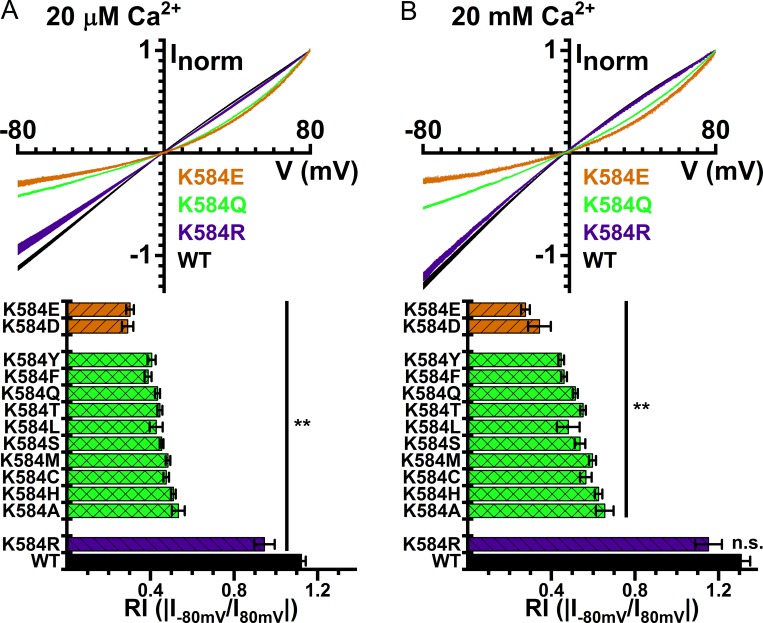
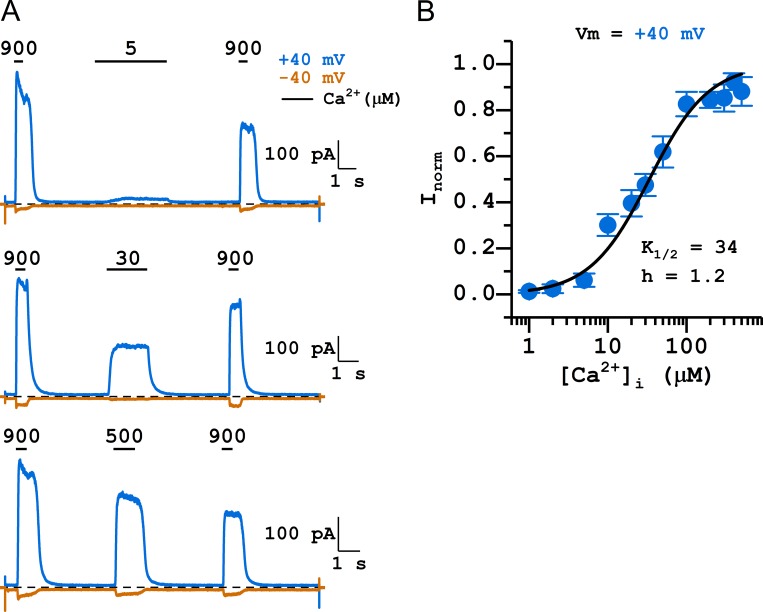
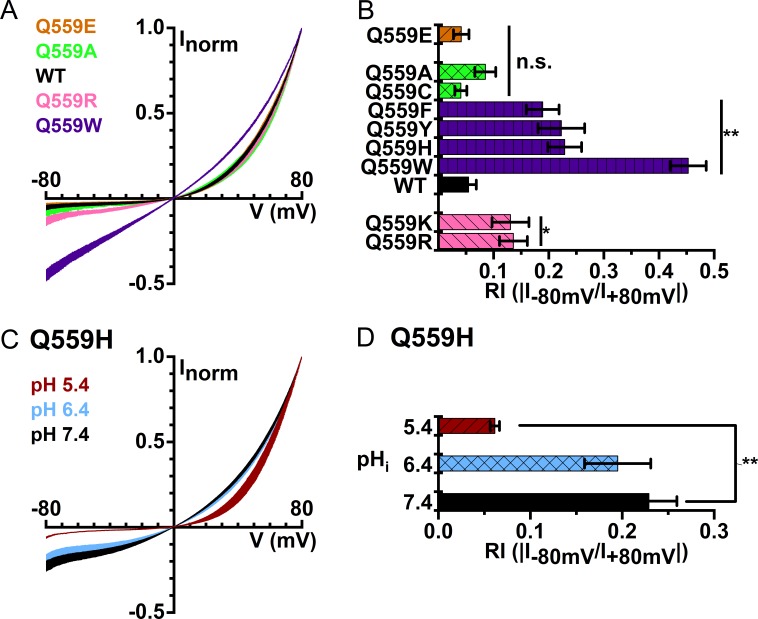
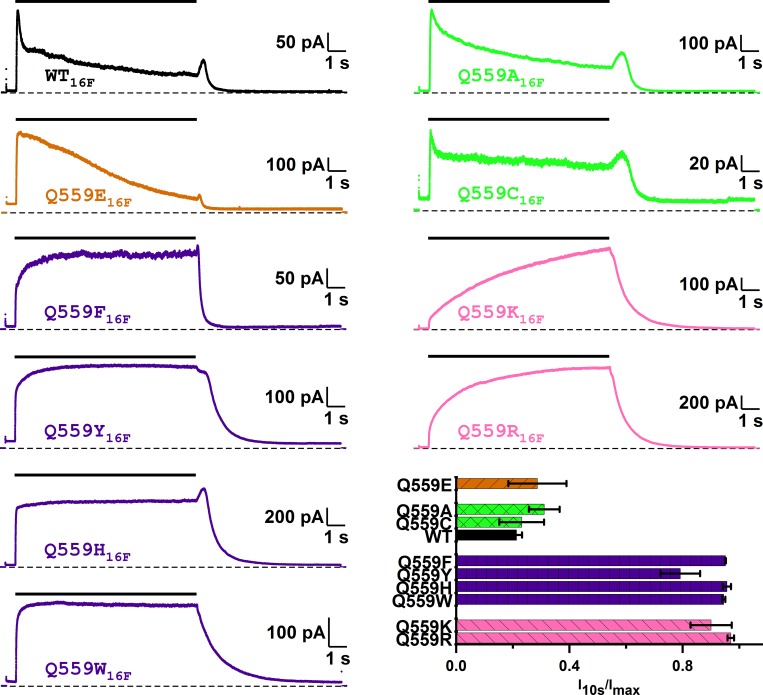
Two TMEM16 family members, TMEM16A and TMEM16F, have different ion transport properties. Upon activation by intracellular Ca, TMEM16A-a Ca-activated Cl channel-is more selective for anions than cations, whereas TMEM16F-a phospholipid scramblase-appears to transport both cations and anions. Under saturating Ca conditions, the current-voltage (I-V) relationships of these two proteins also differ; the I-V curve of TMEM16A is linear, while that of TMEM16F is outwardly rectifying. We previously found that mutating a positively charged lysine residue (K584) in the ion transport pathway to glutamine converted the linear I-V curve of TMEM16A to an outwardly rectifying curve. Interestingly, the corresponding residue in the outwardly rectifying TMEM16F is also a glutamine (Q559). Here, we examine the ion transport functions of TMEM16 molecules and compare the roles of K584 of TMEM16A and Q559 of TMEM16F in controlling the rectification of their respective I-V curves. We find that rectification of TMEM16A is regulated electrostatically by the side-chain charge on the residue at position 584, whereas the charge on residue 559 in TMEM16F has little effect. Unexpectedly, mutation of Q559 to aromatic amino acid residues significantly alters outward rectification in TMEM16F. These same mutants show reduced Ca-induced current rundown (or desensitization) compared with wild-type TMEM16F. A mutant that removes the rundown of TMEM16F could facilitate the study of ion transport mechanisms in this phospholipid scramblase in the same way that a CLC-0 mutant in which inactivation (or closure of the slow gate) is suppressed was used in our previous studies.
TMEM16 家族的两个成员,TMEM16A 和 TMEM16F,具有不同的离子转运特性。在细胞内 Ca2+的激活下,TMEM16A-a Ca2+激活的 Cl-通道-对阴离子的选择性高于阳离子,而 TMEM16F-a 磷脂翻转酶-似乎同时转运阳离子和阴离子。在饱和 Ca2+条件下,这两种蛋白的电流-电压(I-V)关系也不同;TMEM16A 的 I-V 曲线是线性的,而 TMEM16F 的 I-V 曲线是外向整流的。我们之前发现,将离子转运途径中带正电荷的赖氨酸残基(K584)突变为谷氨酰胺,将 TMEM16A 的线性 I-V 曲线转化为外向整流曲线。有趣的是,外向整流的 TMEM16F 中的相应残基也是谷氨酰胺(Q559)。在这里,我们研究了 TMEM16 分子的离子转运功能,并比较了 TMEM16A 的 K584 和 TMEM16F 的 Q559 残基在控制各自 I-V 曲线整流中的作用。我们发现,TMEM16A 的整流是通过位于 584 位的侧链电荷静电调节的,而 TMEM16F 中 559 位残基的电荷几乎没有影响。出乎意料的是,将 Q559 突变为芳香族氨基酸残基会显著改变 TMEM16F 的外向整流。与野生型 TMEM16F 相比,这些相同的突变体显示出 Ca2+诱导的电流衰减(或脱敏)减少。与我们之前的研究中使用的抑制失活(或慢门关闭)的 CLC-0 突变体一样,一种消除 TMEM16F 电流衰减的突变体可以促进对这种磷脂翻转酶离子转运机制的研究。