Department of Biology, Florida International University-Biscayne Bay Campus, North Miami, FL 33181, USA.
BMC Evol Biol. 2013 Jun 20;13:128. doi: 10.1186/1471-2148-13-128.
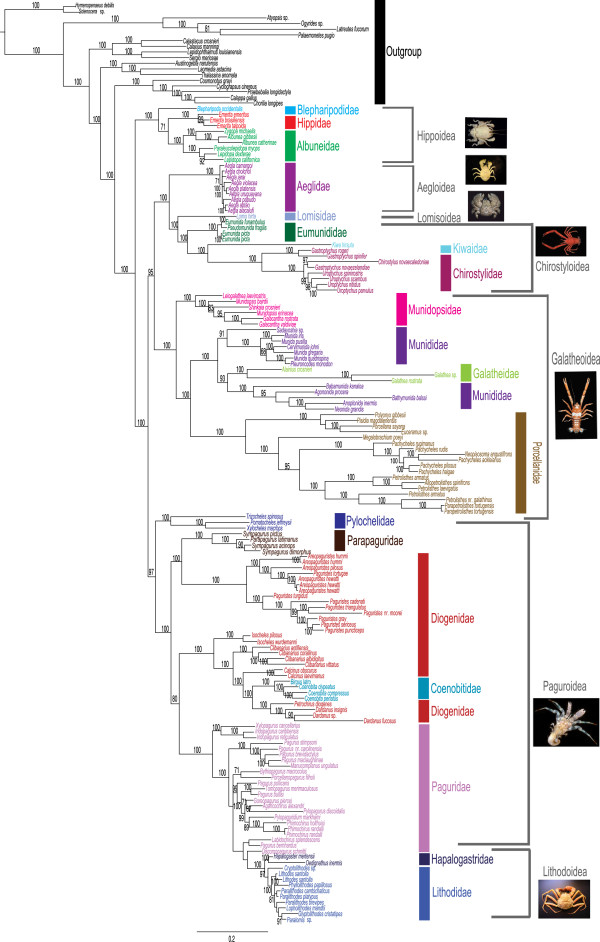
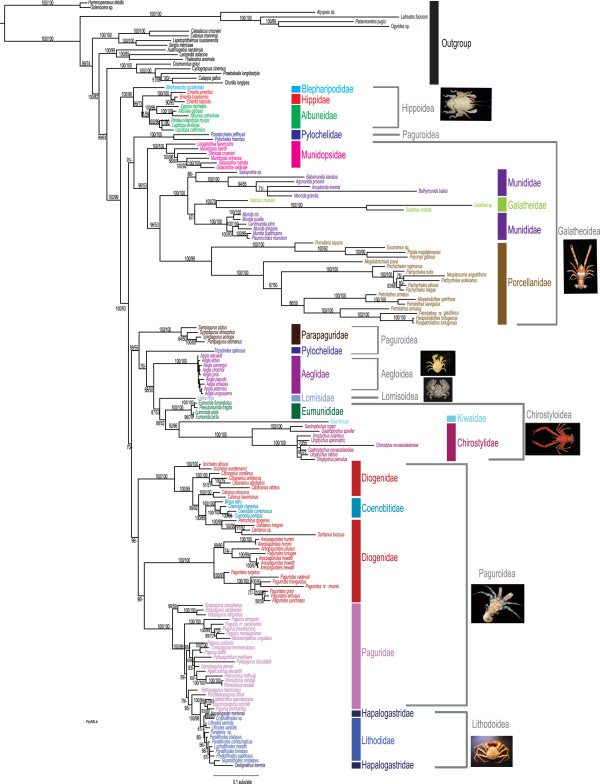
The infraorder Anomura has long captivated the attention of evolutionary biologists due to its impressive morphological diversity and ecological adaptations. To date, 2500 extant species have been described but phylogenetic relationships at high taxonomic levels remain unresolved. Here, we reconstruct the evolutionary history-phylogeny, divergence times, character evolution and diversification-of this speciose clade. For this purpose, we sequenced two mitochondrial (16S and 12S) and three nuclear (H3, 18S and 28S) markers for 19 of the 20 extant families, using traditional Sanger and next-generation 454 sequencing methods. Molecular data were combined with 156 morphological characters in order to estimate the largest anomuran phylogeny to date. The anomuran fossil record allowed us to incorporate 31 fossils for divergence time analyses.
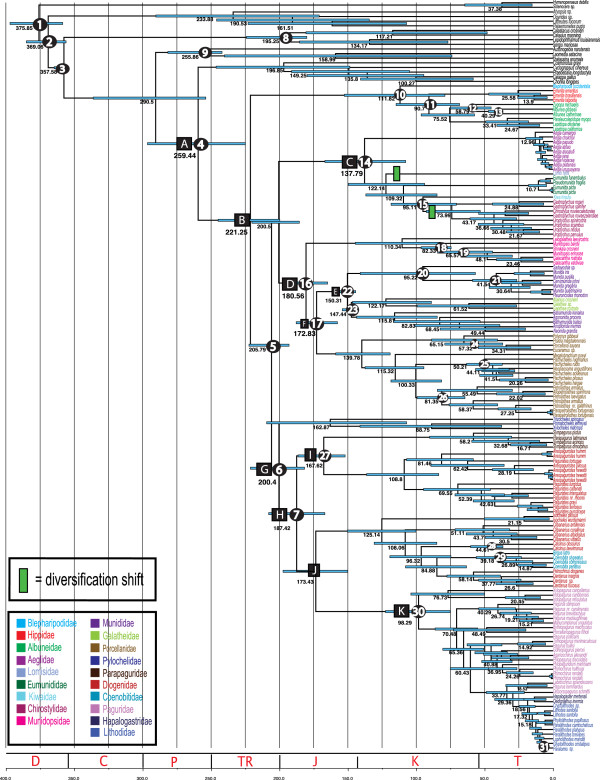
Our best phylogenetic hypothesis (morphological + molecular data) supports most anomuran superfamilies and families as monophyletic. However, three families and eleven genera are recovered as para- and polyphyletic. Divergence time analysis dates the origin of Anomura to the Late Permian ~259 (224-296) MYA with many of the present day families radiating during the Jurassic and Early Cretaceous. Ancestral state reconstruction suggests that carcinization occurred independently 3 times within the group. The invasion of freshwater and terrestrial environments both occurred between the Late Cretaceous and Tertiary. Diversification analyses found the speciation rate to be low across Anomura, and we identify 2 major changes in the tempo of diversification; the most significant at the base of a clade that includes the squat-lobster family Chirostylidae.
Our findings are compared against current classifications and previous hypotheses of anomuran relationships. Many families and genera appear to be poly- or paraphyletic suggesting a need for further taxonomic revisions at these levels. A divergence time analysis provides key insights into the origins of major lineages and events and the timing of morphological (body form) and ecological (habitat) transitions. Living anomuran biodiversity is the product of 2 major changes in the tempo of diversification; our initial insights suggest that the acquisition of a crab-like form did not act as a key innovation.
由于其令人印象深刻的形态多样性和生态适应性,次目(亚目)十足目长期以来一直吸引着进化生物学家的注意。迄今为止,已描述了 2500 种现存物种,但高分类水平的系统发育关系仍未解决。在这里,我们重建了这个物种丰富的分支的进化历史——系统发育、分歧时间、特征进化和多样化。为此,我们使用传统的 Sanger 和下一代 454 测序方法,对 20 个现存科中的 19 个科的两个线粒体(16S 和 12S)和三个核(H3、18S 和 28S)标记进行了测序。为了估计迄今为止最大的十足目系统发育,我们将分子数据与 156 个形态特征相结合。十足目化石记录使我们能够将 31 个化石纳入分歧时间分析。
我们的最佳系统发育假说(形态学+分子数据)支持大多数十足目超科和科是单系的。然而,三个科和十一个属被恢复为并系和多系。分歧时间分析将十足目的起源日期定为晚二叠世~259(224-296)百万年前,许多现代科在侏罗纪和早白垩世辐射。祖先状态重建表明,该组内独立发生了 3 次癌变。淡水和陆地环境的入侵都发生在白垩纪晚期和第三纪之间。多样化分析发现十足目种形成率很低,我们确定了进化速度的 2 个主要变化;最显著的是在包括 squat-lobster 科 Chirostylidae 的一个分支的基部。
我们的研究结果与当前的分类和以前的十足目关系假说进行了比较。许多科和属似乎是并系或多系的,这表明需要在这些水平上进一步进行分类修订。分歧时间分析为主要谱系和事件的起源以及形态(体型)和生态(栖息地)转变的时间提供了关键见解。现存十足目生物多样性是进化速度两次重大变化的产物;我们的初步见解表明,获得类似螃蟹的形态并不是一个关键的创新。