From the MRC National Institute for Medical Research, Mill Hill, London, NW7 1AA, United Kingdom and; Institut für Zelluläre Physiologie and Center for NanoScience, Physiologisches Institut, Ludwig Maximilians Universität, Munich 80336, Germany.
From the MRC National Institute for Medical Research, Mill Hill, London, NW7 1AA, United Kingdom and.
J Biol Chem. 2013 Aug 30;288(35):25183-25193. doi: 10.1074/jbc.M113.484667. Epub 2013 Jul 9.
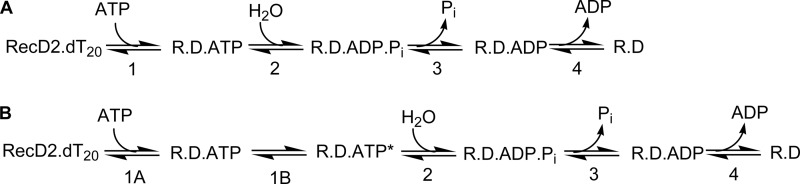
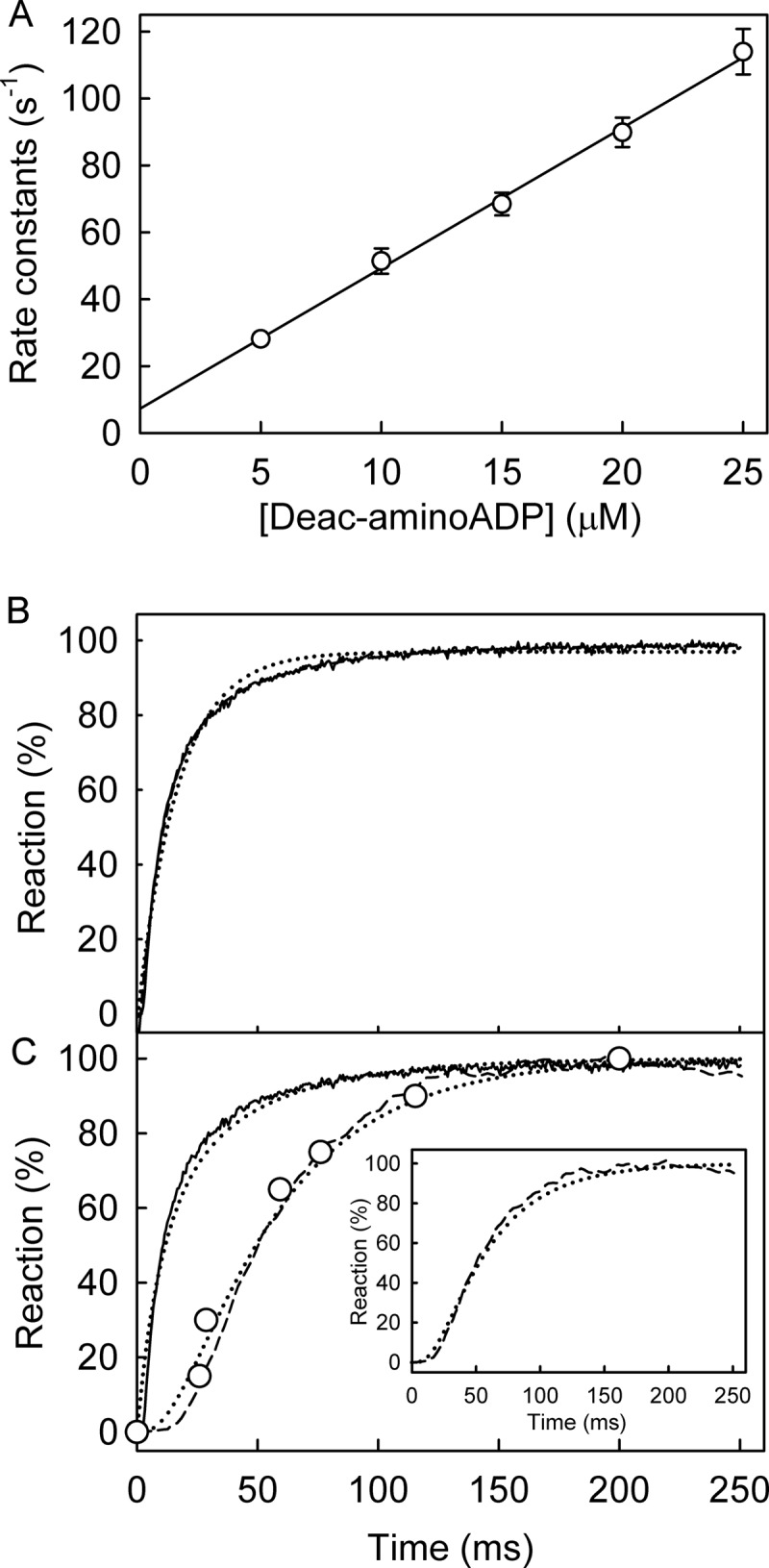
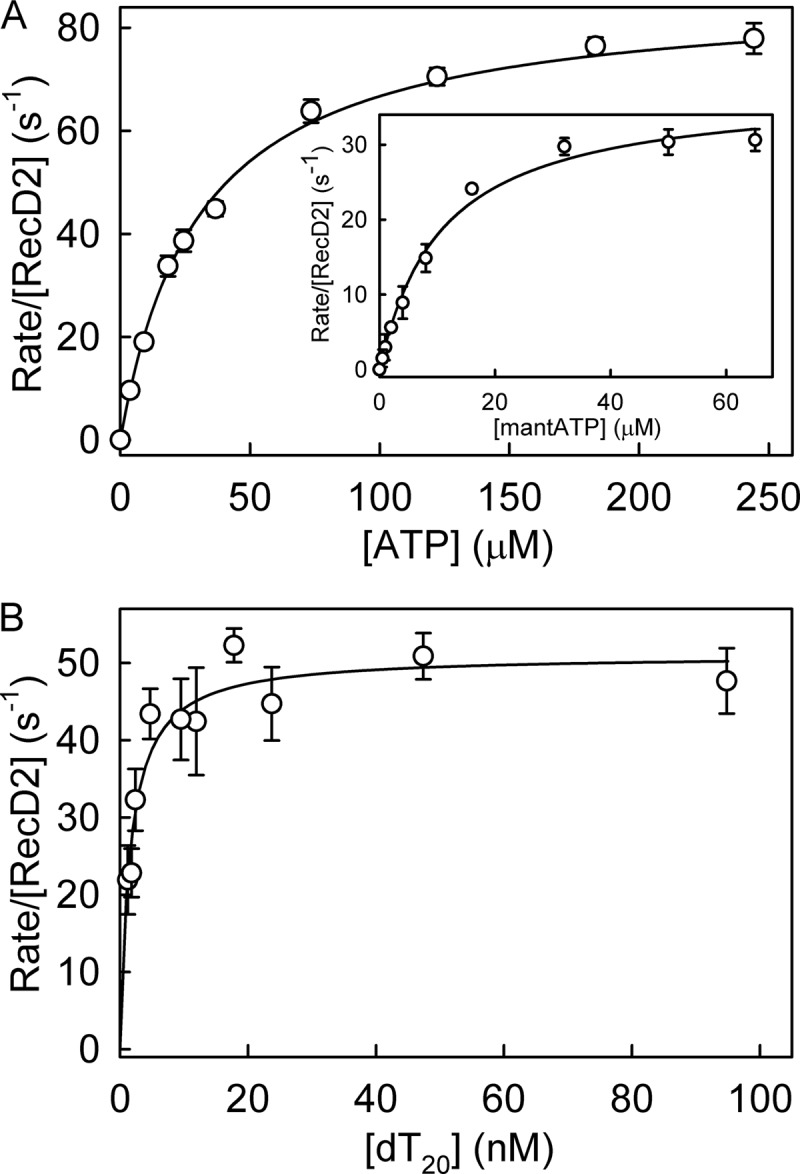
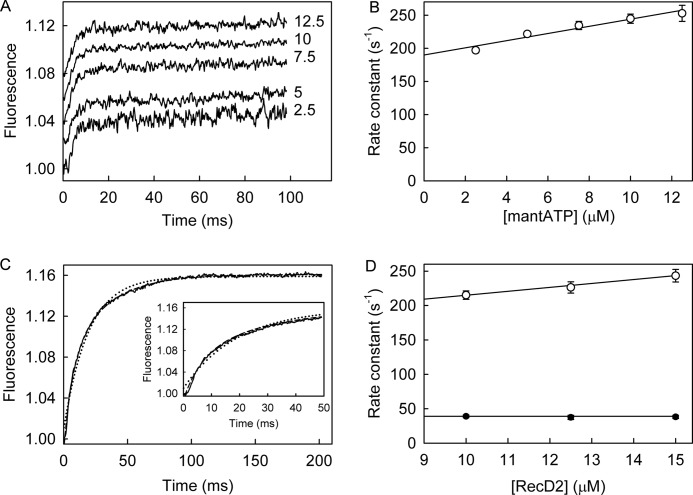
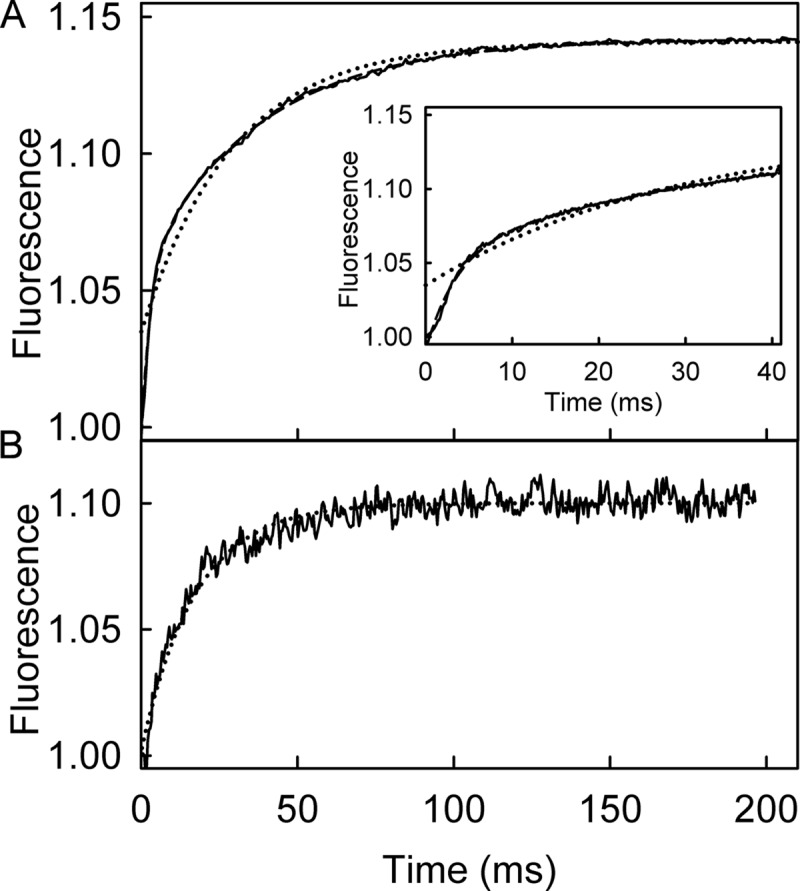
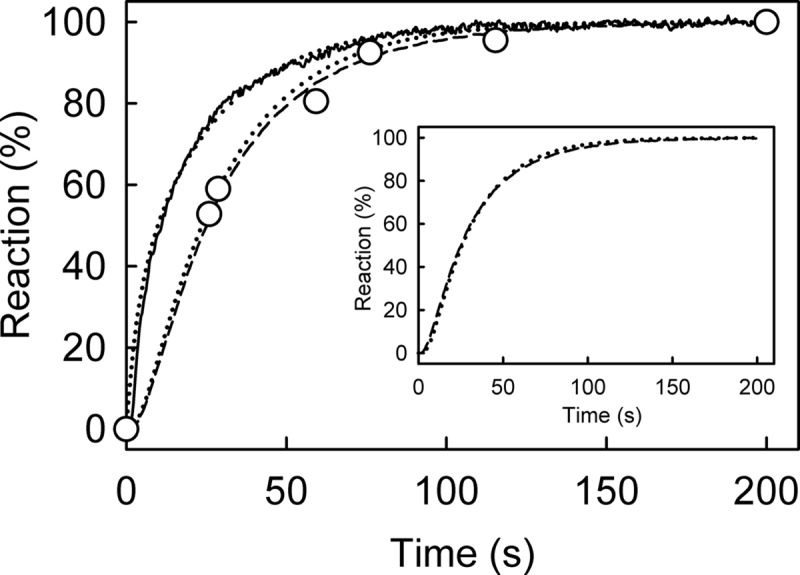
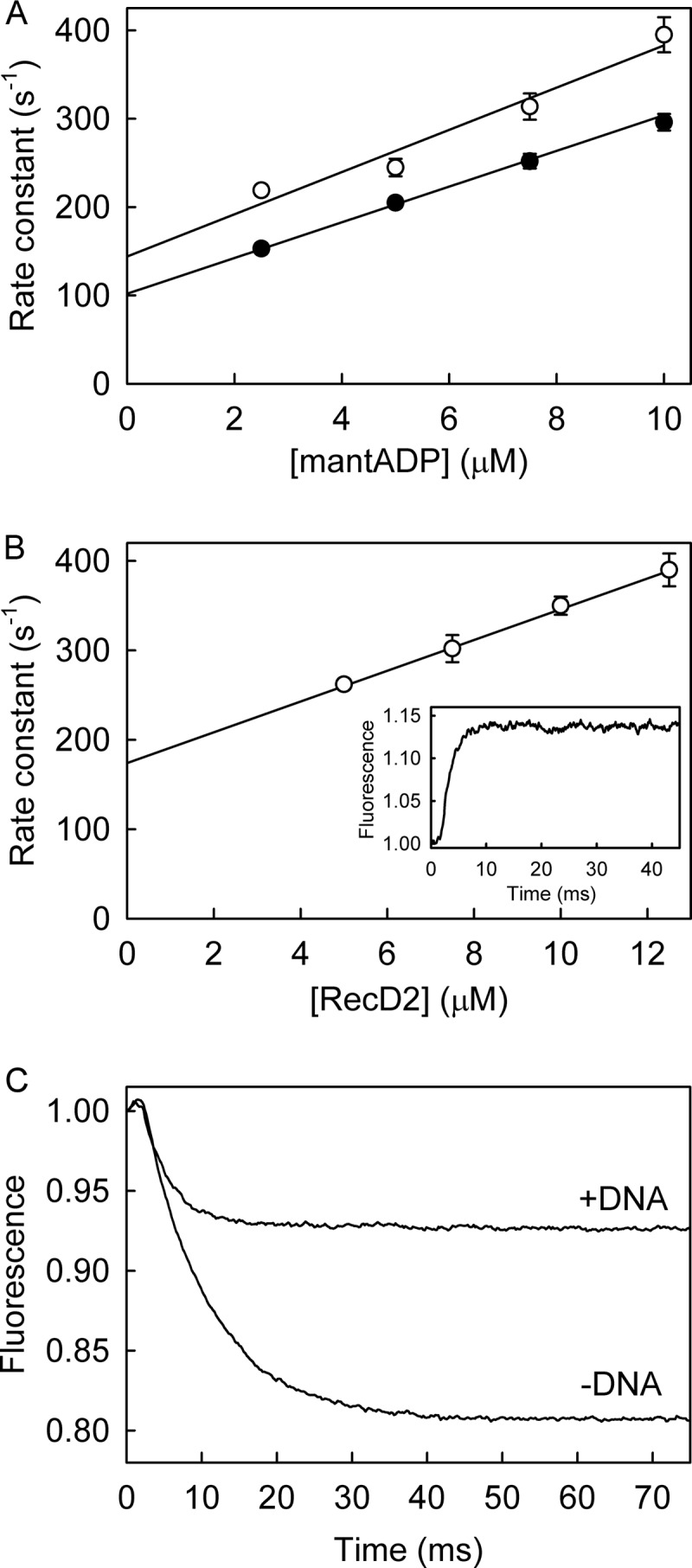
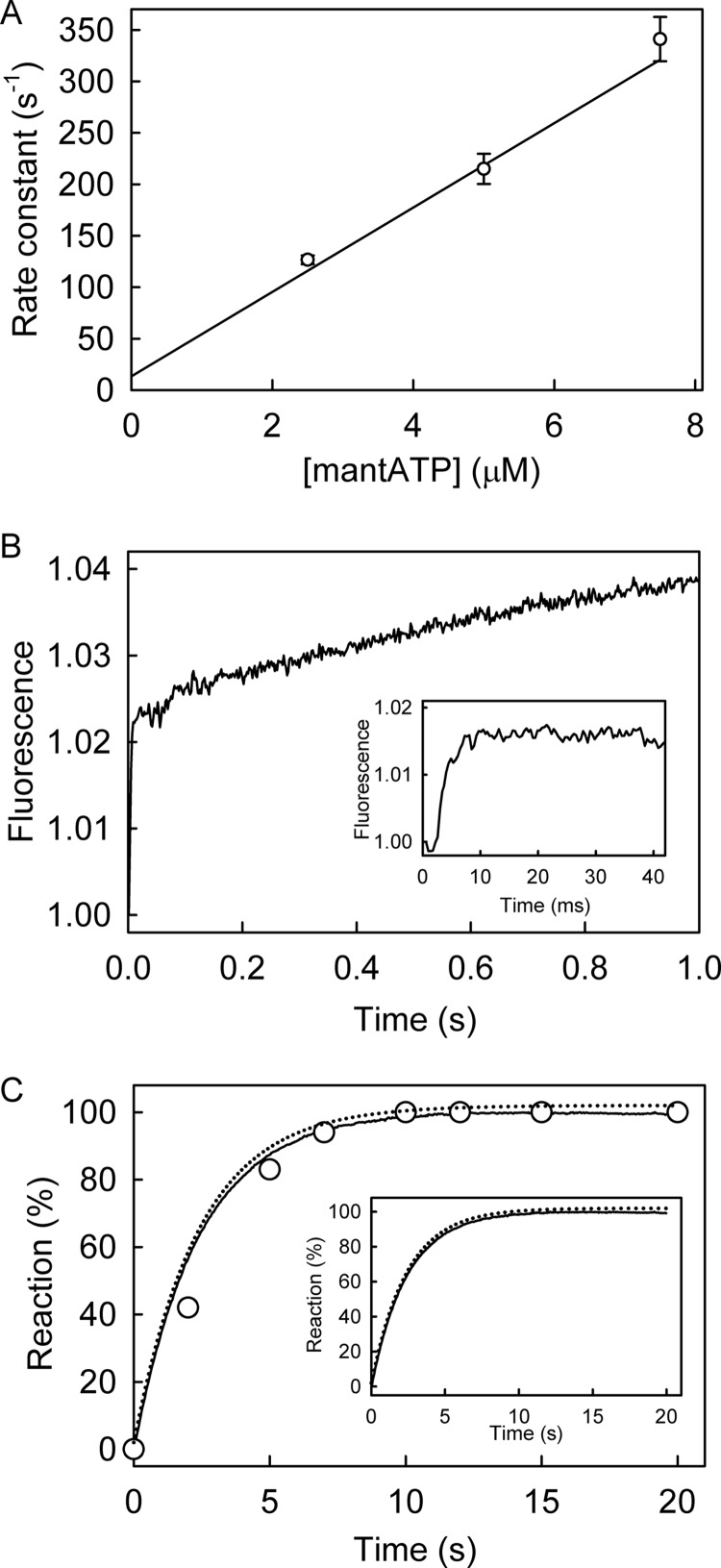
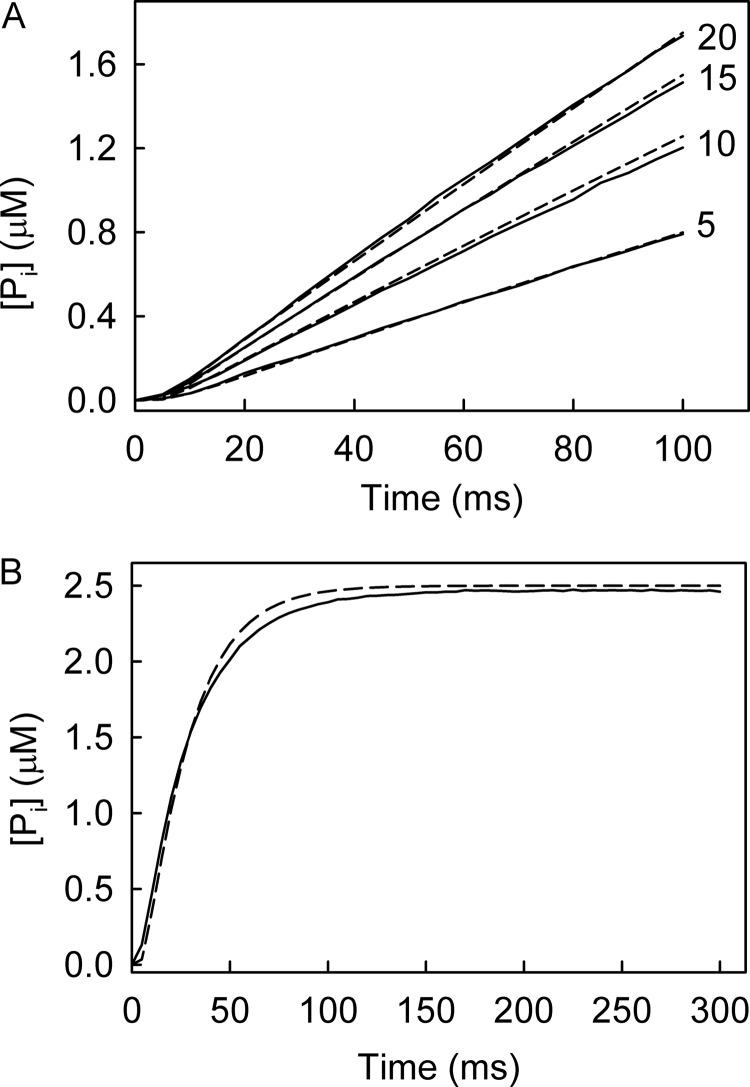
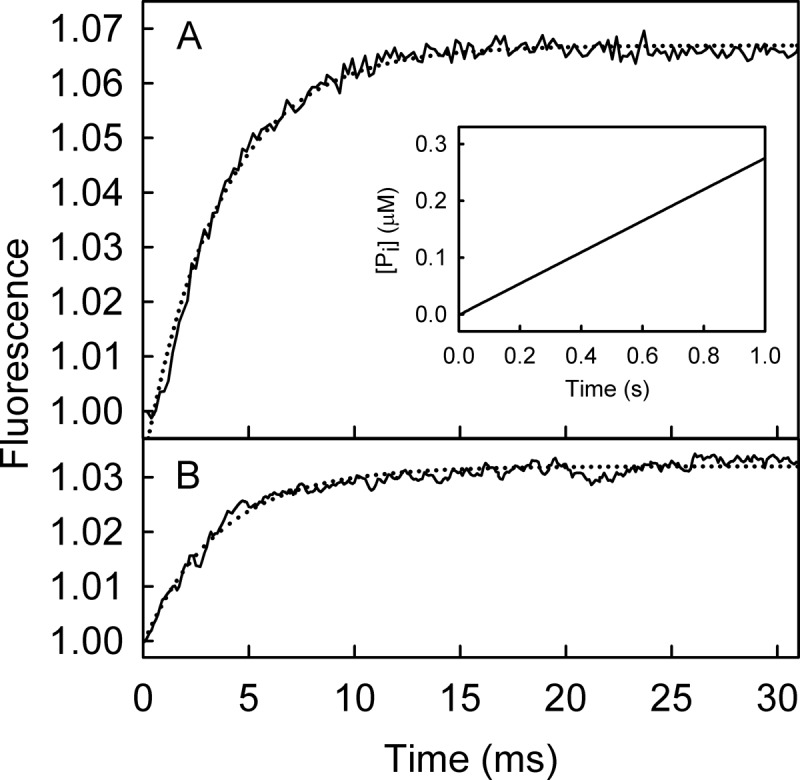
The superfamily 1 helicase, RecD2, is a monomeric, bacterial enzyme with a role in DNA repair, but with 5'-3' activity unlike most enzymes from this superfamily. Rate constants were determined for steps within the ATPase cycle of RecD2 in the presence of ssDNA. The fluorescent ATP analog, mantATP (2'(3')-O-(N-methylanthraniloyl)ATP), was used throughout to provide a complete set of rate constants and determine the mechanism of the cycle for a single nucleotide species. Fluorescence stopped-flow measurements were used to determine rate constants for adenosine nucleotide binding and release, quenched-flow measurements were used for the hydrolytic cleavage step, and the fluorescent phosphate biosensor was used for phosphate release kinetics. Some rate constants could also be measured using the natural substrate, ATP, and these suggested a similar mechanism to that obtained with mantATP. The data show that a rearrangement linked to Mg(2+) coordination, which occurs before the hydrolysis step, is rate-limiting in the cycle and that this step is greatly accelerated by bound DNA. This is also shown here for the PcrA 3'-5' helicase and so may be a general mechanism governing superfamily 1 helicases. The mechanism accounts for the tight coupling between translocation and ATPase activity.
超家族 1 解旋酶 RecD2 是一种单体酶,具有 DNA 修复功能,但与该超家族的大多数酶不同,它具有 5'-3'活性。在 ssDNA 存在的情况下,测定了 RecD2 的 ATP 酶循环中的步骤的速率常数。荧光 ATP 类似物 mantATP(2'(3')-O-(N-甲基蒽酰基)ATP)在整个过程中被用于提供完整的速率常数,并确定单个核苷酸物种的循环机制。荧光停止流动测量用于确定腺苷核苷酸结合和释放的速率常数,猝灭流动测量用于水解裂解步骤,而荧光磷酸生物传感器用于磷酸释放动力学。一些速率常数也可以使用天然底物 ATP 进行测量,这些结果表明与 mantATP 获得的机制相似。数据表明,在水解步骤之前发生的与 Mg(2+)配位相关的重排是循环中的限速步骤,并且结合的 DNA 大大加速了该步骤。这也在 PcrA 3'-5'解旋酶中得到证实,因此可能是一种普遍的机制来控制超家族 1 解旋酶。该机制解释了转位和 ATP 酶活性之间的紧密偶联。