Laboratoire de Physique Statistique, Ecole Normale Superieure, UPMC Univ Paris 06, Universit Paris Diderot, CNRS, 24 rue Lhomond, 75005 Paris, France.
Nucleic Acids Res. 2010 Sep;38(16):5518-26. doi: 10.1093/nar/gkq273. Epub 2010 Apr 27.
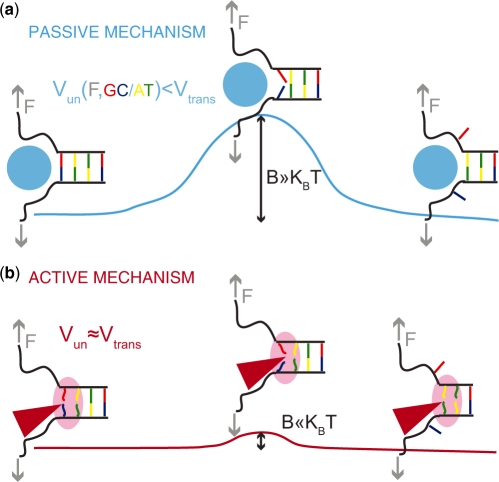
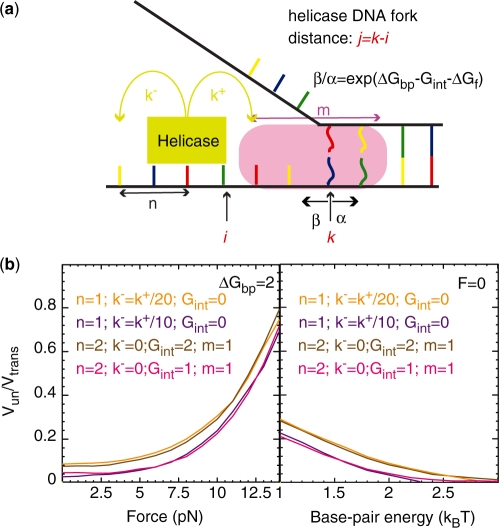
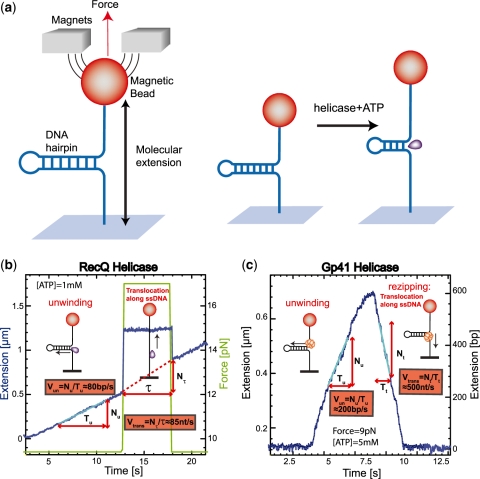
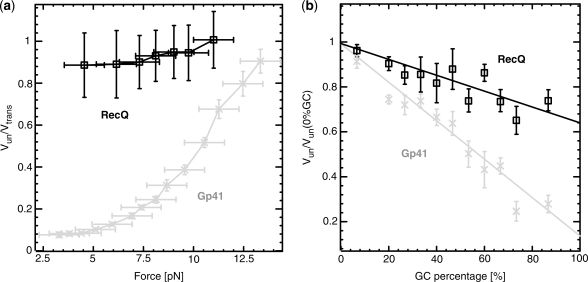
In this work, we discuss the active or passive character of helicases. In the past years, several studies have used the theoretical framework proposed by Betterton and Julicher [Betterton, M.D. and Julicher, F. (2005) Opening of nucleic-acid double strands by helicases: active versus passive opening. Phys. Rev. E, 71, 11904-11911.] to analyse the unwinding data and assess the mechanism of the helicase under study (active versus passive). However, this procedure has given rise to apparently contradictory interpretations: helicases exhibiting similar behaviour have been classified as both active and passive enzymes [Johnson, D.S., Bai, L. Smith, B.Y., Patel, S.S. and Wang, M.D. (2007) Single-molecule studies reveal dynamics of DNA unwinding by the ring-shaped T7 helicase. Cell, 129, 1299-1309; Lionnet, T., Spiering, M.M., Benkovic, S.J., Bensimon, D. and Croquette, V. (2007) Real-time observation of bacteriophage T4 gp41 helicase reveals an unwinding mechanism Proc. Natl Acid. Sci., 104, 19790-19795]. In this work, we show that when the helicase under study has not been previously well characterized (namely, if its step size and rate of slippage are unknown) a multi-parameter fit to the afore-mentioned model can indeed lead to contradictory interpretations. We thus propose to differentiate between active and passive helicases on the basis of the comparison between their observed translocation velocity on single-stranded nucleic acid and their unwinding rate of double-stranded nucleic acid (with various GC content and under different tensions). A threshold separating active from passive behaviour is proposed following an analysis of the reported activities of different helicases. We study and contrast the mechanism of two helicases that exemplify these two behaviours: active for the RecQ helicase and passive for the gp41 helicase.
在这项工作中,我们讨论了解旋酶的主动或被动特征。在过去的几年中,已有几项研究使用了 Betterton 和 Julicher [Betterton, M.D. and Julicher, F. (2005) Opening of nucleic-acid double strands by helicases: active versus passive opening. Phys. Rev. E, 71, 11904-11911.]提出的理论框架来分析解旋数据,并评估所研究的解旋酶的机制(主动或被动)。然而,这一过程产生了明显矛盾的解释:表现出相似行为的解旋酶被归类为主动和被动酶[Johnson, D.S., Bai, L. Smith, B.Y., Patel, S.S. and Wang, M.D. (2007) Single-molecule studies reveal dynamics of DNA unwinding by the ring-shaped T7 helicase. Cell, 129, 1299-1309; Lionnet, T., Spiering, M.M., Benkovic, S.J., Bensimon, D. and Croquette, V. (2007) Real-time observation of bacteriophage T4 gp41 helicase reveals an unwinding mechanism Proc. Natl Acid. Sci., 104, 19790-19795]。在这项工作中,我们表明,当所研究的解旋酶以前没有得到很好的表征时(即,如果其步幅和滑移率未知),对上述模型的多参数拟合确实可能导致矛盾的解释。因此,我们建议根据观察到的单链核酸上的迁移速度与双链核酸的解旋速度(具有不同的 GC 含量和不同的张力)之间的比较,将主动和被动解旋酶区分开来。在分析了不同解旋酶的报道活性之后,提出了一个区分主动和被动行为的阈值。我们研究并对比了两种解旋酶的机制,它们分别代表了这两种行为:RecQ 解旋酶为主动型,gp41 解旋酶为被动型。