Graduate Program in Genetics and Genomics, University of Florida Genetics Institute, Gainesville, Florida, United States of America.
PLoS One. 2013 Jun 28;8(6):e67156. doi: 10.1371/journal.pone.0067156. Print 2013.
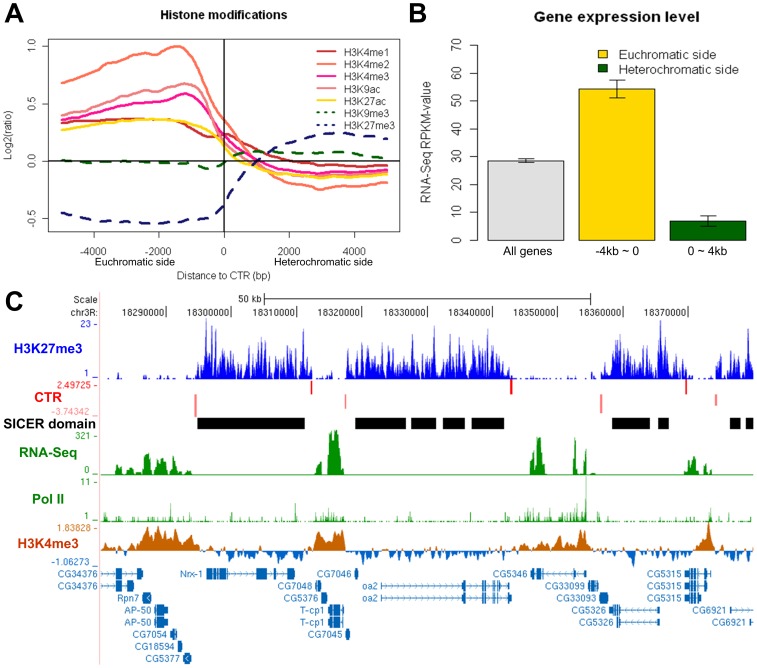
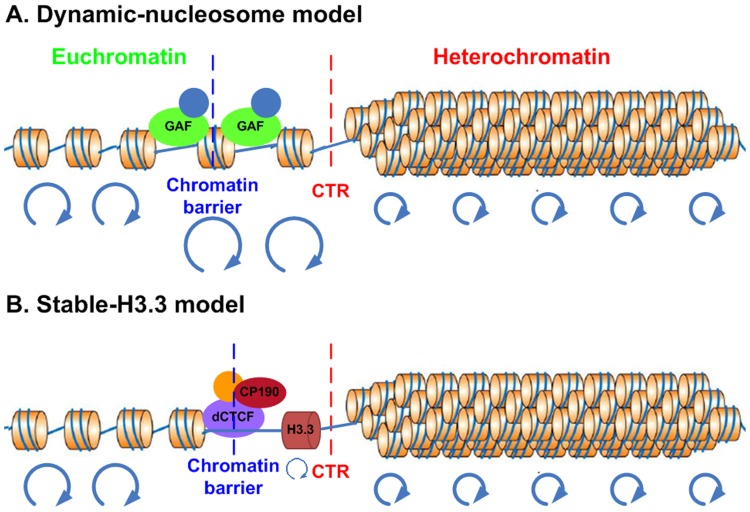
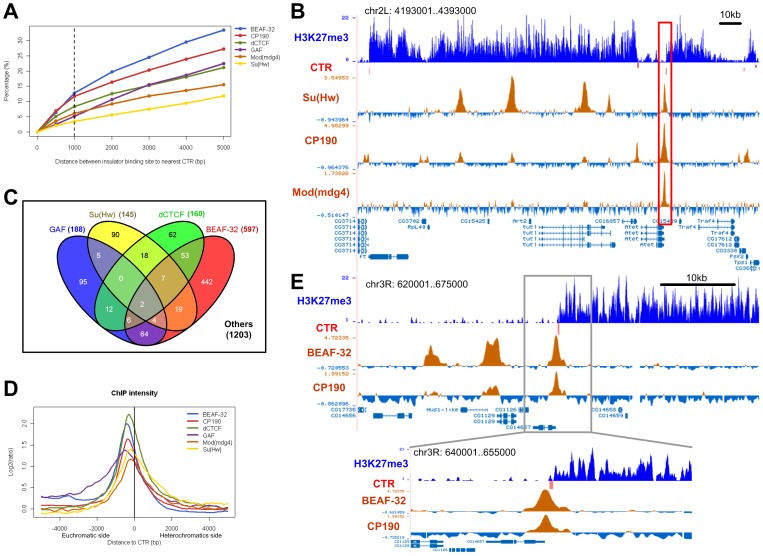
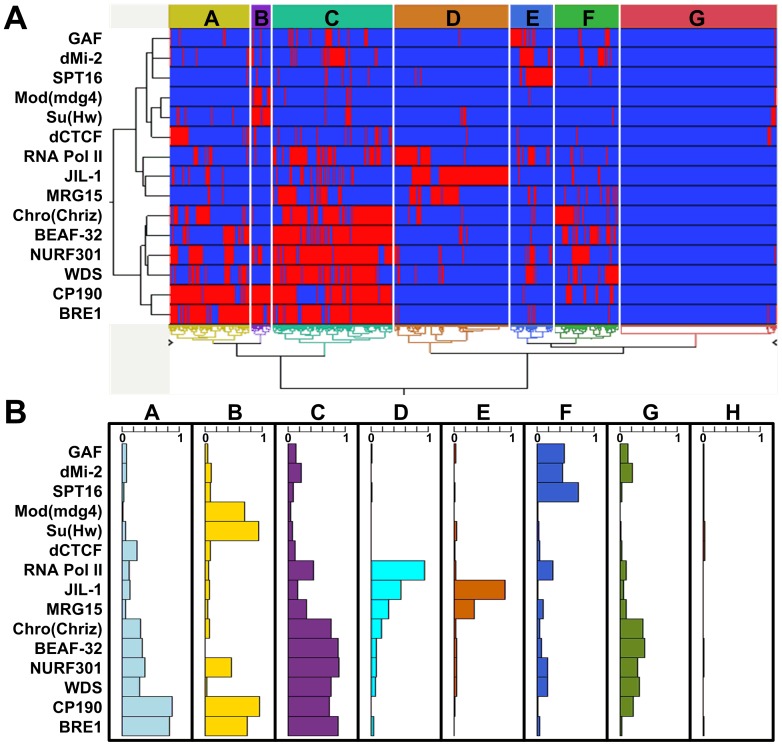
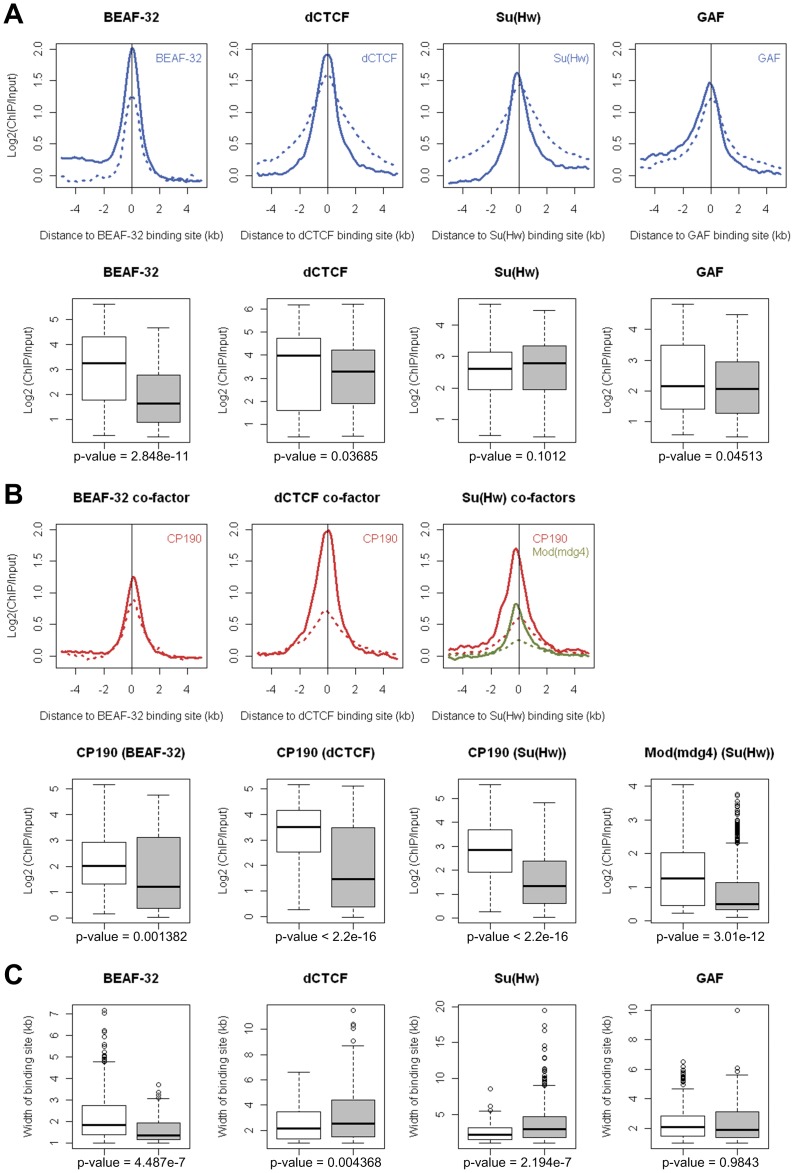
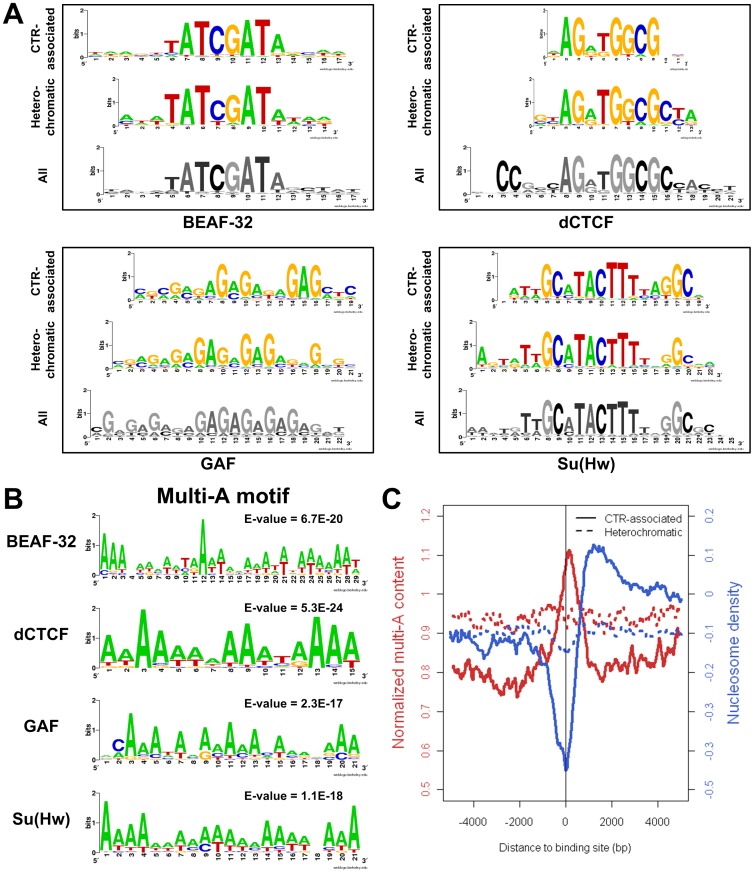
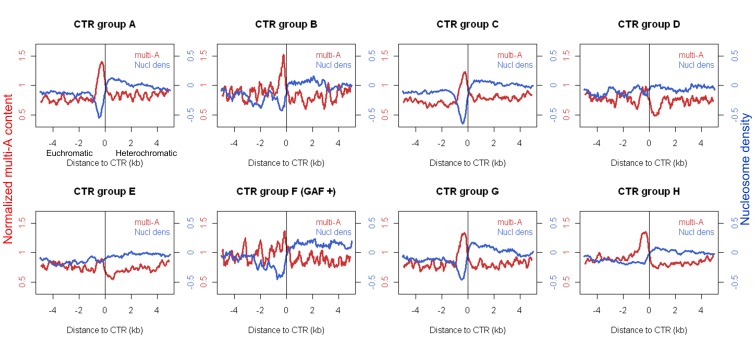
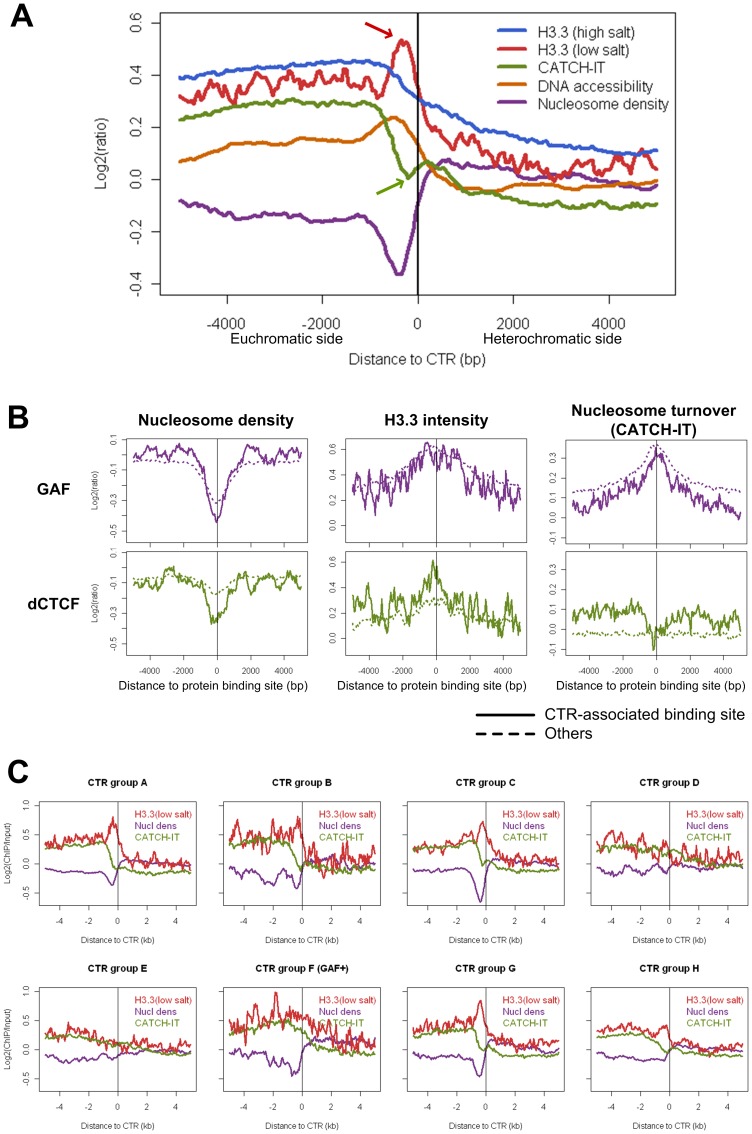
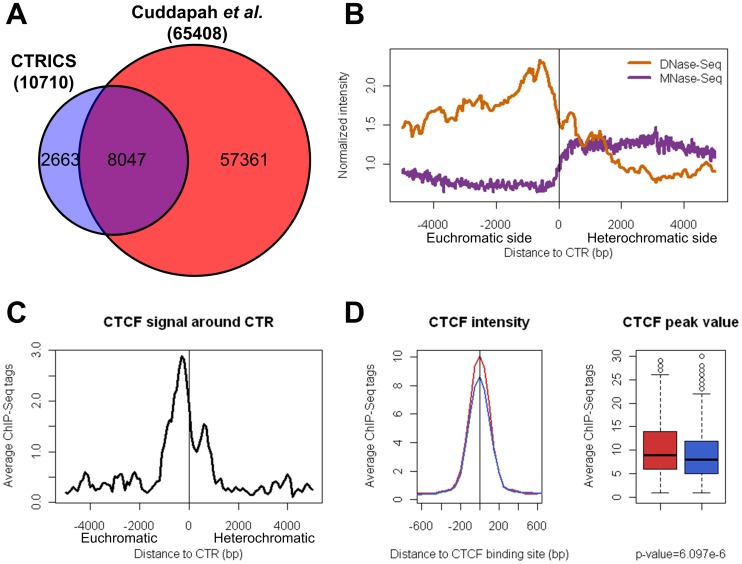
Due to the self-propagating nature of the heterochromatic modification H3K27me3, chromatin barrier activities are required to demarcate the boundary and prevent it from encroaching into euchromatic regions. Studies in Drosophila and vertebrate systems have revealed several important chromatin barrier elements and their respective binding factors. However, epigenomic data indicate that the binding of these factors are not exclusive to chromatin boundaries. To gain a comprehensive understanding of facultative heterochromatin boundaries, we developed a two-tiered method to identify the Chromatin Transitional Region (CTR), i.e. the nucleosomal region that shows the greatest transition rate of the H3K27me3 modification as revealed by ChIP-Seq. This approach was applied to identify CTRs in Drosophila S2 cells and human HeLa cells. Although many insulator proteins have been characterized in Drosophila, less than half of the CTRs in S2 cells are associated with known insulator proteins, indicating unknown mechanisms remain to be characterized. Our analysis also revealed that the peak binding of insulator proteins are usually 1-2 nucleosomes away from the CTR. Comparison of CTR-associated insulator protein binding sites vs. those in heterochromatic region revealed that boundary-associated binding sites are distinctively flanked by nucleosome destabilizing sequences, which correlates with significant decreased nucleosome density and increased binding intensities of co-factors. Interestingly, several subgroups of boundaries have enhanced H3.3 incorporation but reduced nucleosome turnover rate. Our genome-wide study reveals that diverse mechanisms are employed to define the boundaries of facultative heterochromatin. In both Drosophila and mammalian systems, only a small fraction of insulator protein binding sites co-localize with H3K27me3 boundaries. However, boundary-associated insulator binding sites are distinctively flanked by nucleosome destabilizing sequences, which correlates with significantly decreased nucleosome density and increased binding of co-factors.
由于异染色质修饰 H3K27me3 的自我传播性质,染色质屏障活性需要对边界进行划分,以防止其侵入常染色质区域。在果蝇和脊椎动物系统中的研究揭示了几个重要的染色质屏障元件及其各自的结合因子。然而,表观基因组数据表明,这些因子的结合并不局限于染色质边界。为了全面了解可塑异染色质边界,我们开发了一种两级方法来识别染色质过渡区(CTR),即在 ChIP-Seq 揭示的 H3K27me3 修饰中表现出最大转变率的核小体区域。该方法应用于鉴定果蝇 S2 细胞和人 HeLa 细胞中的 CTR。尽管在果蝇中已经鉴定了许多绝缘子蛋白,但 S2 细胞中只有不到一半的 CTR 与已知的绝缘子蛋白相关,这表明仍有未知的机制有待研究。我们的分析还表明,绝缘子蛋白的峰结合通常距离 CTR 有 1-2 个核小体。将 CTR 相关绝缘子蛋白结合位点与异染色质区域的结合位点进行比较,发现边界相关的结合位点周围明显存在核小体不稳定序列,这与核小体密度显著降低和共同因子结合强度增加相关。有趣的是,几个边界亚群的 H3.3 掺入增强,但核小体周转率降低。我们的全基因组研究表明,多种机制被用于定义可塑异染色质的边界。在果蝇和哺乳动物系统中,只有一小部分绝缘子蛋白结合位点与 H3K27me3 边界共定位。然而,边界相关的绝缘子结合位点周围明显存在核小体不稳定序列,这与核小体密度显著降低和共同因子结合强度增加相关。