Department für Integrative Zoologie, Universität Wien, Althanstr, 14, 1090, Wien, Austria.
Front Zool. 2013 Sep 6;10(1):53. doi: 10.1186/1742-9994-10-53.
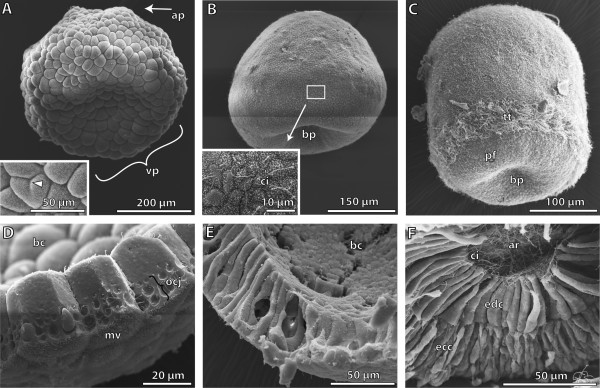
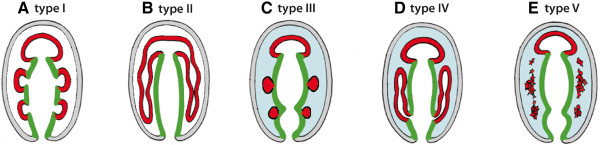
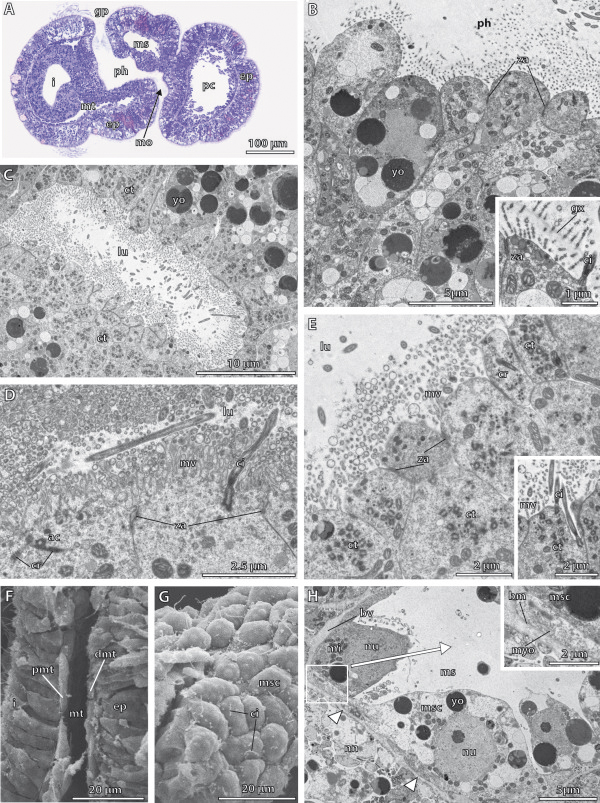
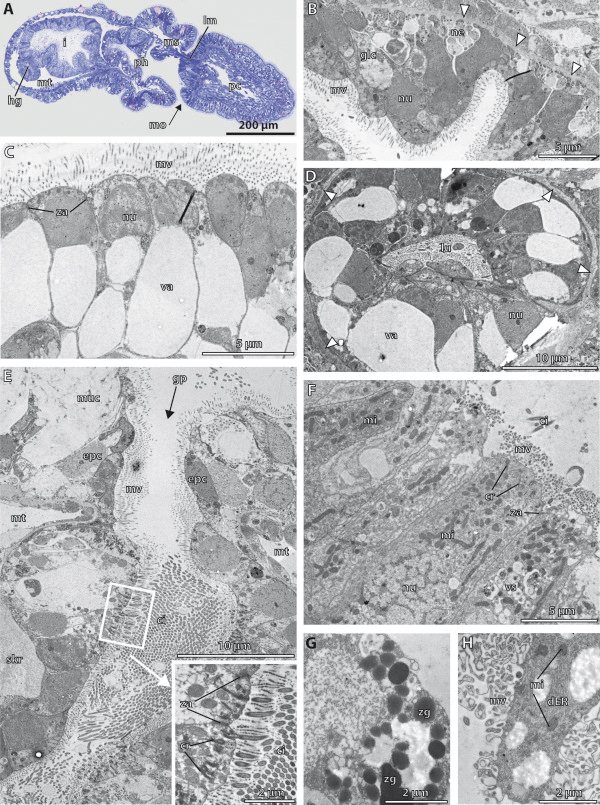
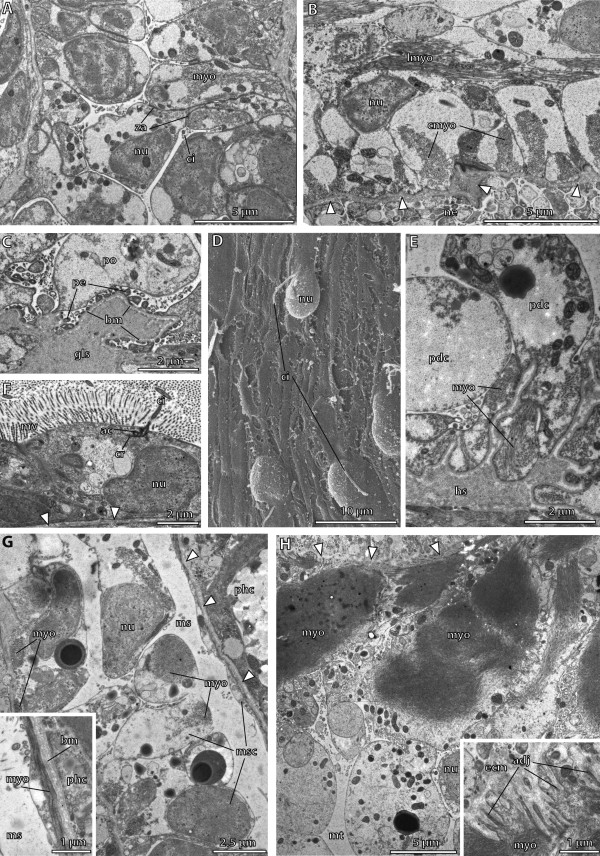
Traditionally, the origin of the third germ layer and its special formation of coelomic cavities by enterocoely is regarded to be an informative character in phylogenetic analyses. In early deuterostomes such as sea urchins, the mesoderm forms through a single evagination pinching off from the apical end of the archenteron which then gives off mesocoela and metacoela on each side. This echinoid-type coelom formation has conventionally been assumed to be ancestral for Deuterostomia. However, recent phylogenetic analyses show that Echinodermata hold a more derived position within Deuterostomia. In this regard a subgroup of Hemichordata, namely enteropneusts, seem to host promising candidates, because they are supposed to have retained many ancestral deuterostome features on the one hand, and furthermore share some characteristics with chordates on the other hand. In enteropneusts a wide range of different modes of coelom formation has been reported and in many cases authors of the original observations carefully detailed the limitations of their descriptions, while these doubts disappeared in subsequent reviews. In the present study, we investigated the development of all tissues in an enteropneust, Saccoglossus kowalevskii by using modern morphological techniques such as complete serial sectioning for LM and TEM, and 3D-reconstructions, in order to contribute new data to the elucidation of deuterostome evolution.
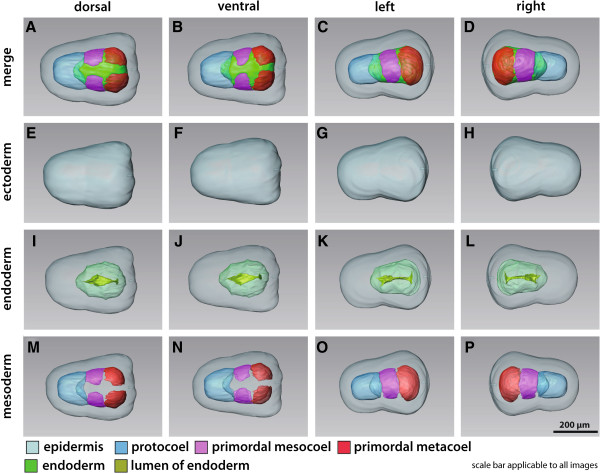
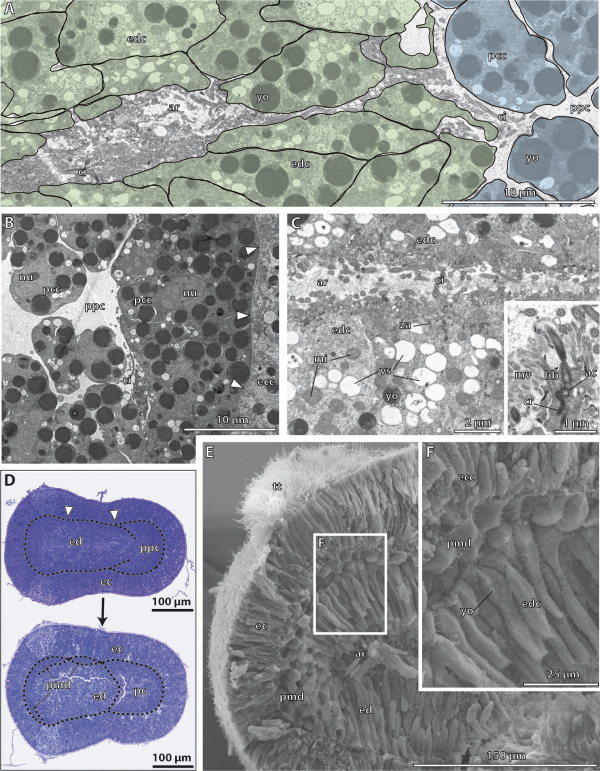
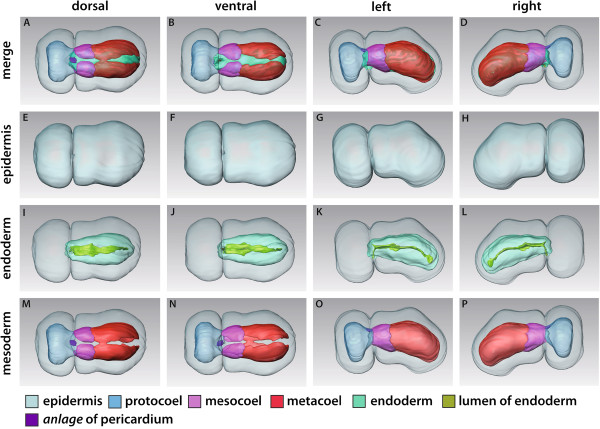
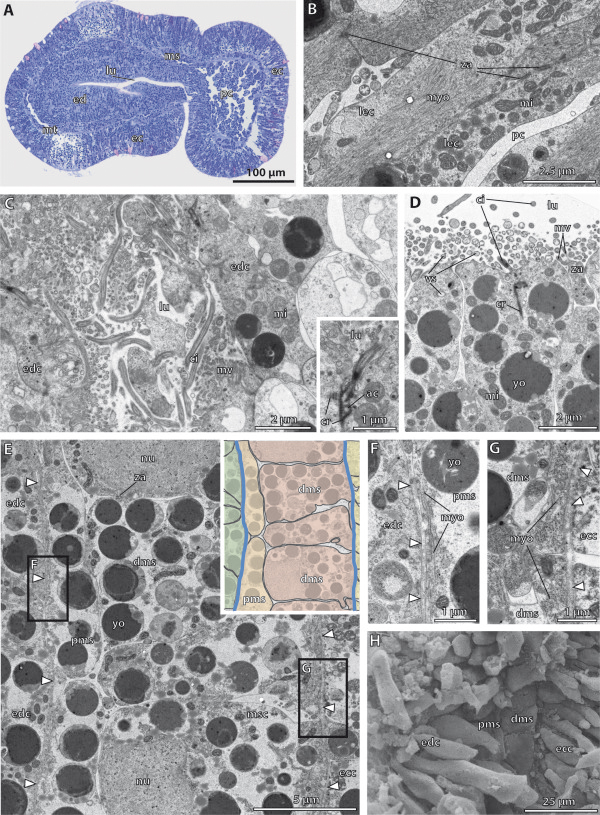
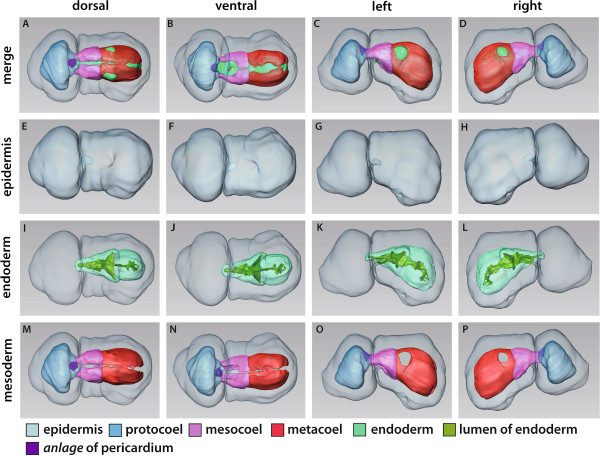
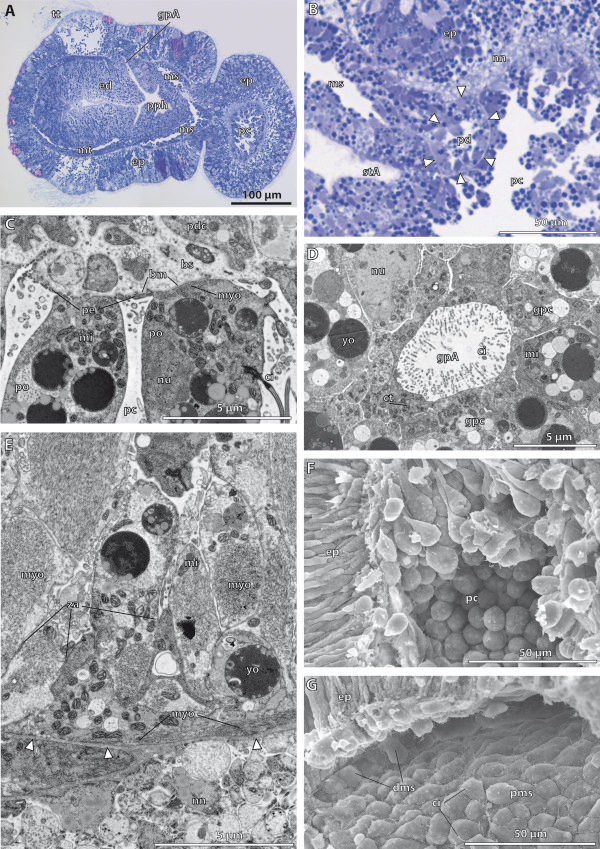
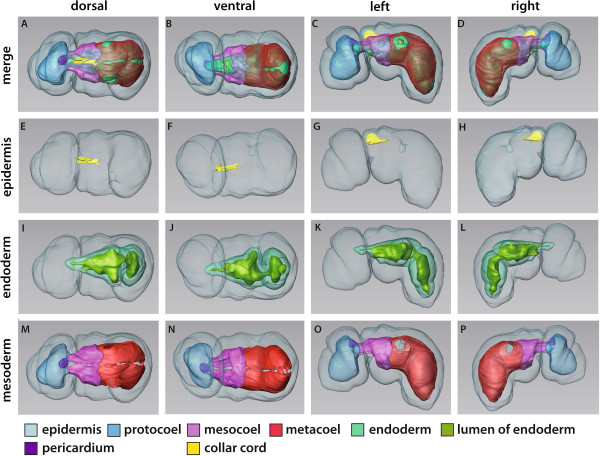
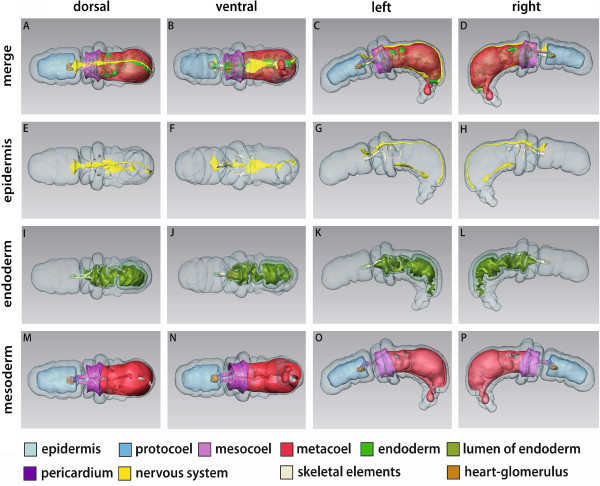
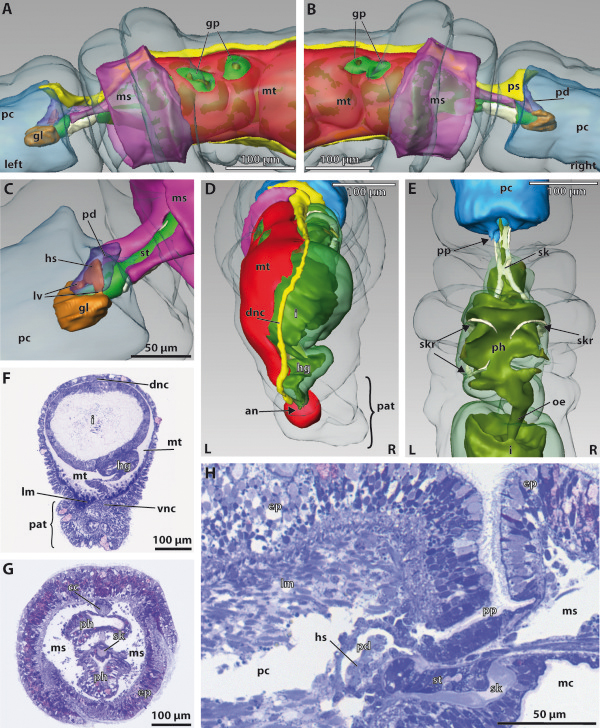
Our data show that in the enteropneust S. kowalevskii all main coelomic cavities (single protocoel, paired mesocoela and metacoela) derive from the endoderm via enterocoely as separate evaginations, in contrast to the aforementioned echinoid-type. The anlagen of the first pair of gill slits emerge at the late kink stage (~96 h pf). From that time onwards, we documented a temporal left-first development of the gill slits and skeletal gill rods in S. kowalevskii until the 2 gill slit juvenile stage.
The condition of coelom formation from separate evaginations is recapitulated in the larva of amphioxus and can be observed in crinoid echinoderms in a similar way. Therefore, coelom formation from separated pouches, rather than from a single apical pouch with eventual subdivision is suggested as the ancestral type of coelom formation for Deuterostomia. Left-right asymmetries are also present in echinoderms (rudiment formation), cephalochordates (larval development), tunicates (gut coiling) and vertebrates (visceral organs), and it is known from other studies applying molecular genetic analyses that genes such as nodal, lefty and pitx are involved during development. We discuss our findings in S. kowalevskii in the light of morphological as well as molecular genetic data.
传统上,第三胚层的起源及其通过肠腔形成体腔的特殊方式被认为是系统发育分析中的一个信息特征。在早期的后口动物如海胆中,中胚层通过从原肠胚的顶端单一的膨出形成,然后在每一侧发出中胚层和后中胚层。这种海胆型体腔形成方式通常被认为是后口动物的祖征。然而,最近的系统发育分析表明,棘皮动物在后口动物中处于更衍生的位置。在这方面,半索动物的一个亚组,即肠鳃动物,似乎是有希望的候选者,因为一方面它们被认为保留了许多原始后口动物的特征,另一方面它们与脊索动物共享一些特征。在肠鳃动物中,已经报道了广泛的不同的体腔形成模式,并且在许多情况下,原始观察的作者详细说明了他们描述的局限性,而这些疑虑在随后的综述中消失了。在本研究中,我们通过使用现代形态学技术,如完整的连续切片用于 LM 和 TEM,以及 3D 重建,研究了肠鳃动物 Saccoglossus kowalevskii 的所有组织的发育,以提供新的数据来阐明后口动物的进化。
我们的数据表明,在肠鳃动物 S. kowalevskii 中,所有主要的体腔腔(单一的原腔、成对的中胚层和后中胚层)都是通过肠腔从内胚层单独形成的,与上述的海胆型不同。第一对鳃裂的原基在晚期扭结阶段(~96 hpf)出现。从那时起,我们记录了 S. kowalevskii 中鳃裂和骨骼鳃棒的左先发育,直到 2 个鳃裂幼体阶段。
分离的囊泡形成体腔的形成方式在文昌鱼的幼虫中得到了重现,并且在棘皮动物的海百合类中也可以以类似的方式观察到。因此,从分离的囊泡而不是从一个具有最终细分的单一顶端囊泡形成体腔被认为是后口动物体腔形成的祖征类型。左右不对称也存在于棘皮动物(原基形成)、头索动物(幼虫发育)、被囊动物(肠道卷曲)和脊椎动物(内脏器官)中,并且从应用分子遗传分析的其他研究中可知,节点、左侧和 pitx 等基因在发育过程中参与其中。我们根据形态学和分子遗传数据讨论了我们在 S. kowalevskii 中的发现。