Institute of Biotechnology, Vilnius University, Graičiūno 8, Vilnius LT-02241, Lithuania.
Nucleic Acids Res. 2014 Feb;42(3):1393-413. doi: 10.1093/nar/gkt900. Epub 2013 Oct 7.
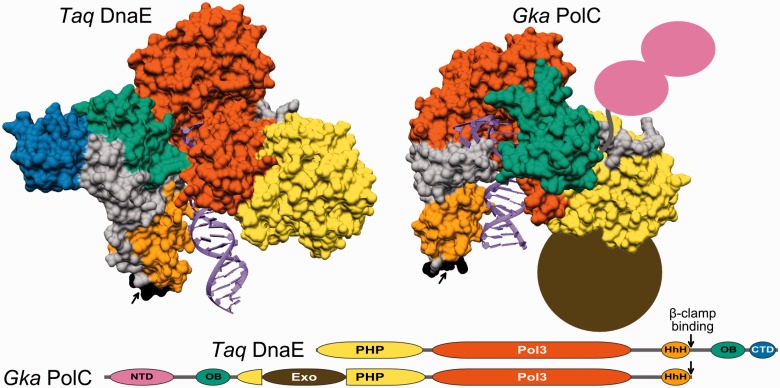
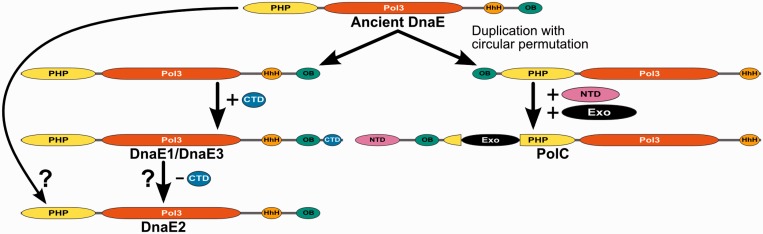
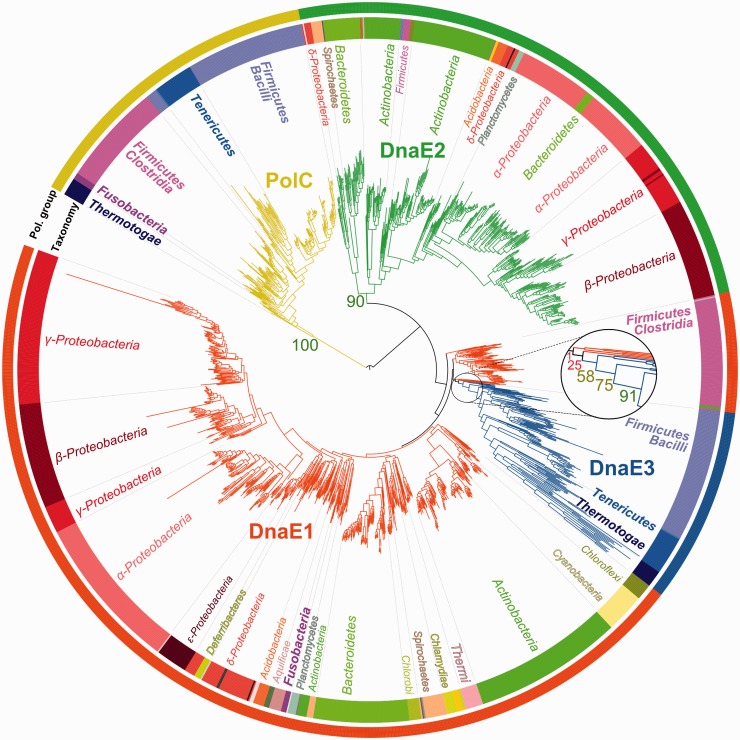
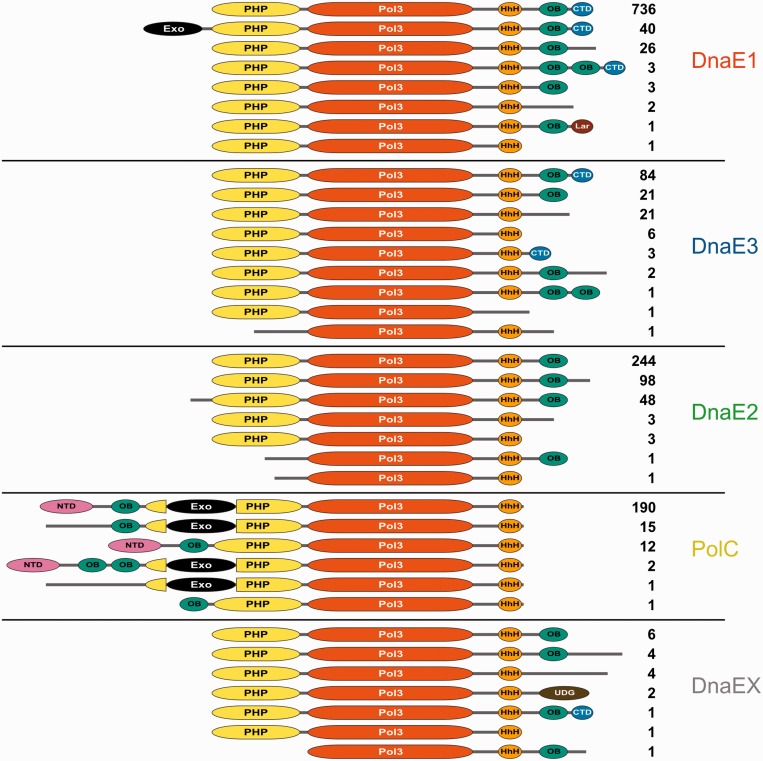
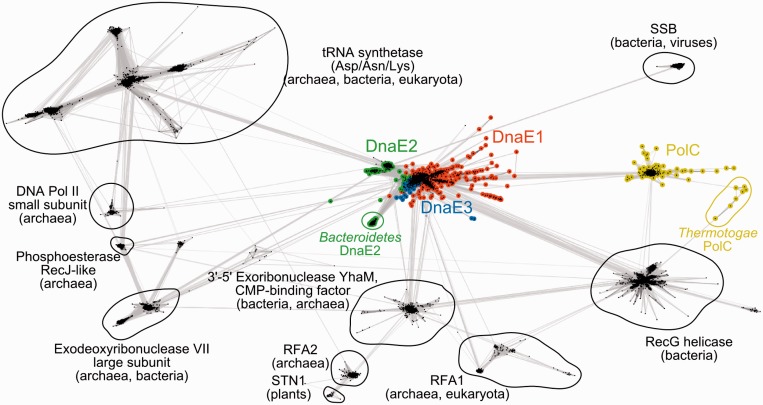
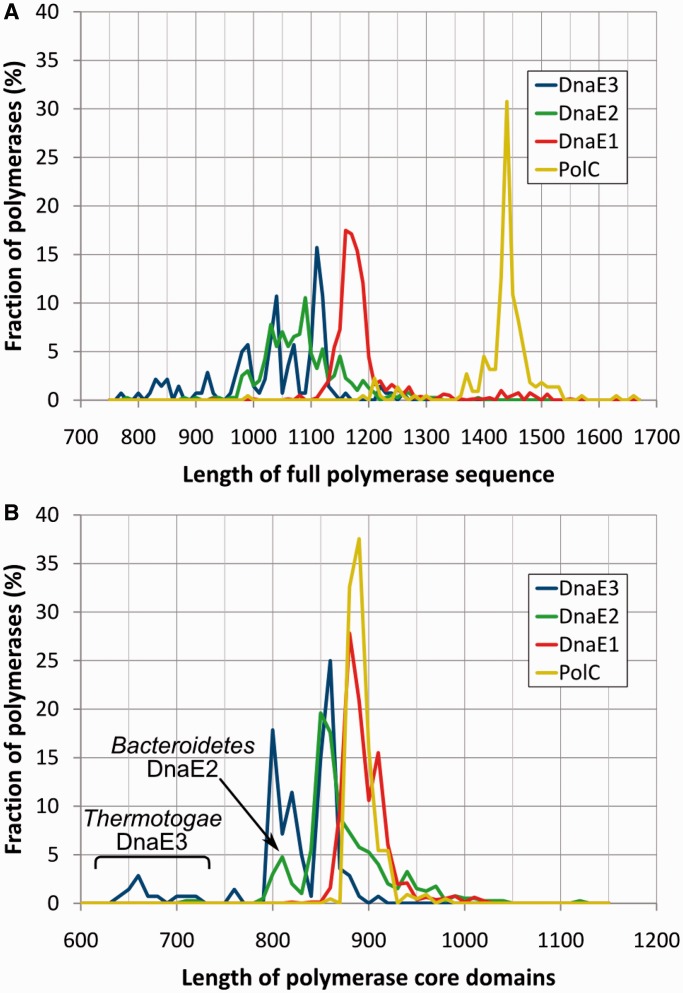
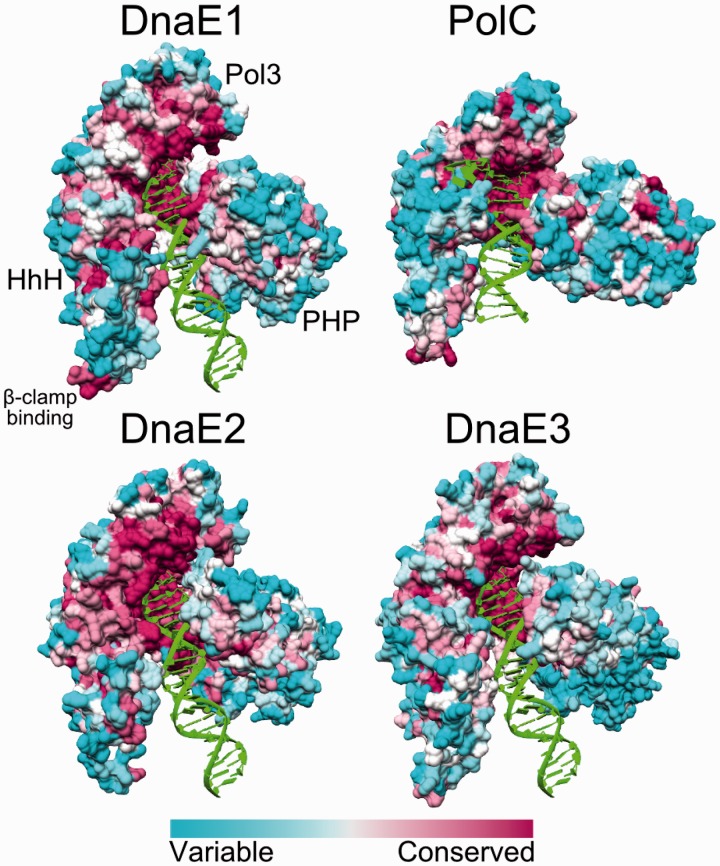
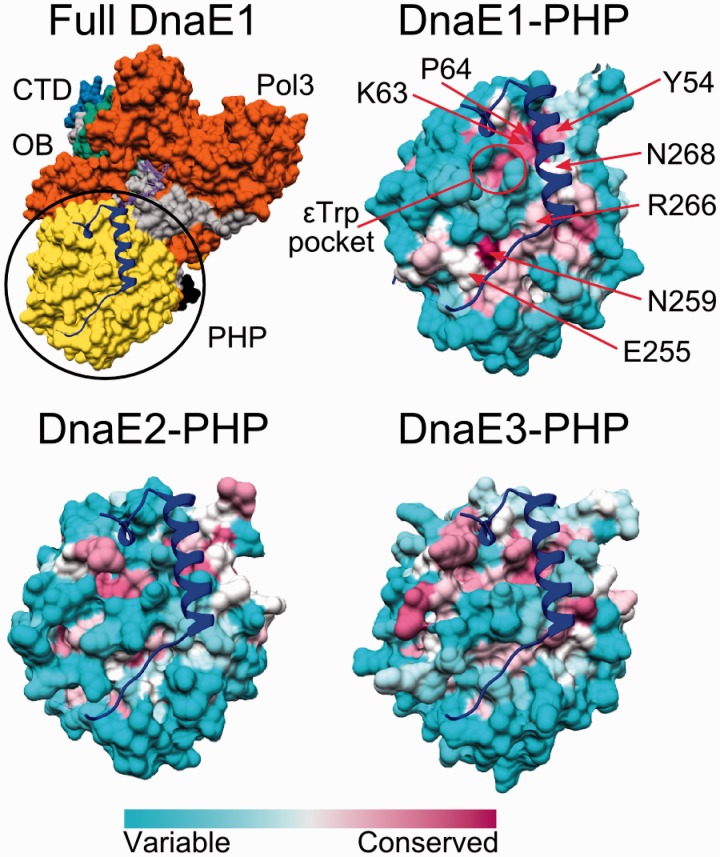
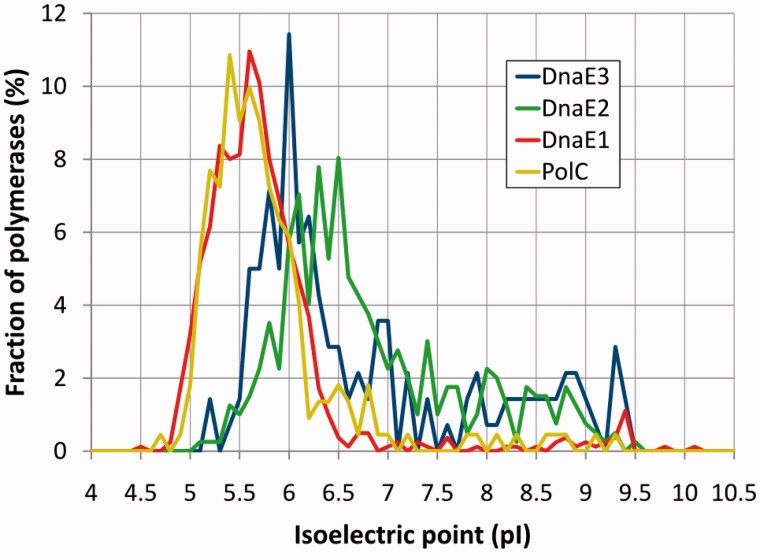
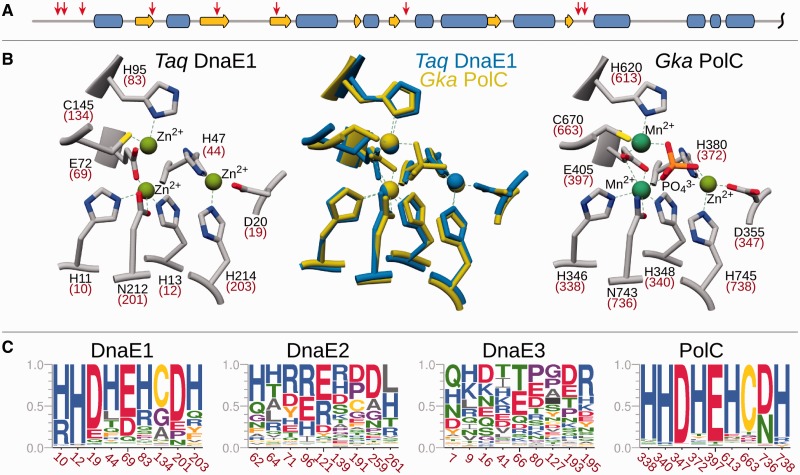
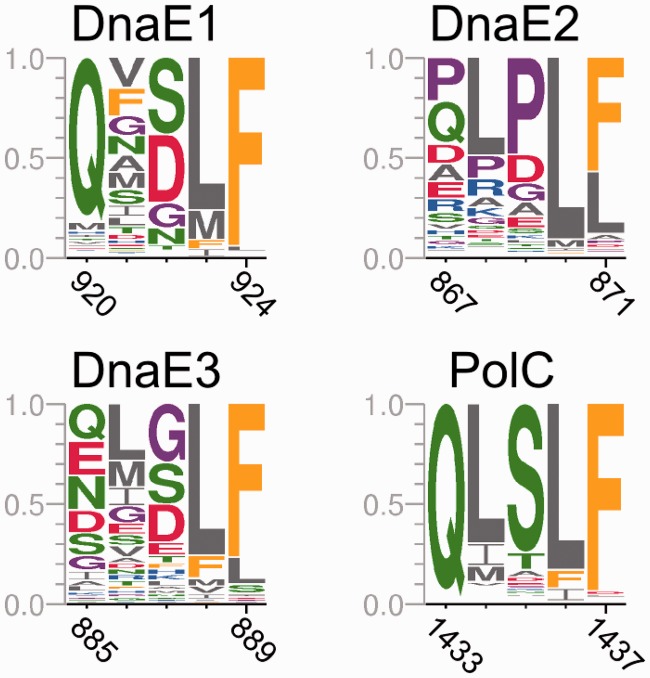
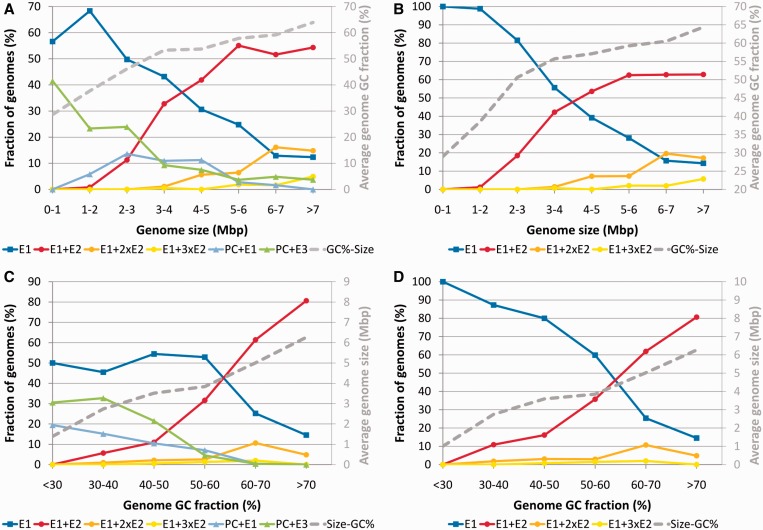
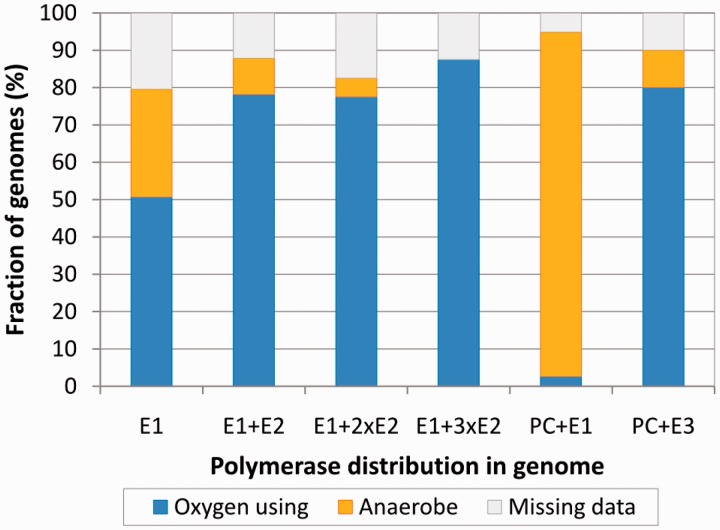
The analysis of ∼ 2000 bacterial genomes revealed that they all, without a single exception, encode one or more DNA polymerase III α-subunit (PolIIIα) homologs. Classified into C-family of DNA polymerases they come in two major forms, PolC and DnaE, related by ancient duplication. While PolC represents an evolutionary compact group, DnaE can be further subdivided into at least three groups (DnaE1-3). We performed an extensive analysis of various sequence, structure and surface properties of all four polymerase groups. Our analysis suggests a specific evolutionary pathway leading to PolC and DnaE from the last common ancestor and reveals important differences between extant polymerase groups. Among them, DnaE1 and PolC show the highest conservation of the analyzed properties. DnaE3 polymerases apparently represent an 'impaired' version of DnaE1. Nonessential DnaE2 polymerases, typical for oxygen-using bacteria with large GC-rich genomes, have a number of features in common with DnaE3 polymerases. The analysis of polymerase distribution in genomes revealed three major combinations: DnaE1 either alone or accompanied by one or more DnaE2s, PolC + DnaE3 and PolC + DnaE1. The first two combinations are present in Escherichia coli and Bacillus subtilis, respectively. The third one (PolC + DnaE1), found in Clostridia, represents a novel, so far experimentally uncharacterized, set.
对约 2000 个细菌基因组的分析表明,它们无一例外地编码一个或多个 DNA 聚合酶 III α 亚基(PolIIIα)同源物。根据它们在 DNA 聚合酶 C 家族中的分类,有两种主要形式,PolC 和 DnaE,它们是由古老的复制而来。虽然 PolC 代表了一个进化紧凑的群体,但 DnaE 可以进一步细分为至少三个群体(DnaE1-3)。我们对所有四种聚合酶群体的各种序列、结构和表面特性进行了广泛的分析。我们的分析表明,从最后一个共同祖先开始,存在一条特定的进化途径,导致 PolC 和 DnaE 的出现,并揭示了现存聚合酶群体之间的重要差异。其中,DnaE1 和 PolC 显示出分析特性的最高保守性。DnaE3 聚合酶显然代表了 DnaE1 的“受损”版本。非必需的 DnaE2 聚合酶,对于具有大 GC 丰富基因组的需氧细菌来说是典型的,与 DnaE3 聚合酶有许多共同的特征。聚合酶在基因组中的分布分析揭示了三种主要的组合:DnaE1 单独存在或与一个或多个 DnaE2 一起存在,PolC + DnaE3 和 PolC + DnaE1。前两种组合分别存在于大肠杆菌和枯草芽孢杆菌中。第三种组合(PolC + DnaE1),在梭菌中发现,代表了一种新的、迄今为止尚未通过实验进行表征的组合。