James D Watson Institute of Genome Sciences, Zhejiang University, Hangzhou 310007, China.
Biol Direct. 2012 Jan 10;7:2. doi: 10.1186/1745-6150-7-2.
As a key parameter of genome sequence variation, the GC content of bacterial genomes has been investigated for over half a century, and many hypotheses have been put forward to explain this GC content variation and its relationship to other fundamental processes. Previously, we classified eubacteria into dnaE-based groups (the dimeric combination of DNA polymerase III alpha subunits), according to a hypothesis where GC content variation is essentially governed by genome replication and DNA repair mechanisms. Further investigation led to the discovery that two major mutator genes, polC and dnaE2, may be responsible for genomic GC content variation. Consequently, an in-depth analysis was conducted to evaluate various potential intrinsic and extrinsic factors in association with GC content variation among eubacterial genomes.
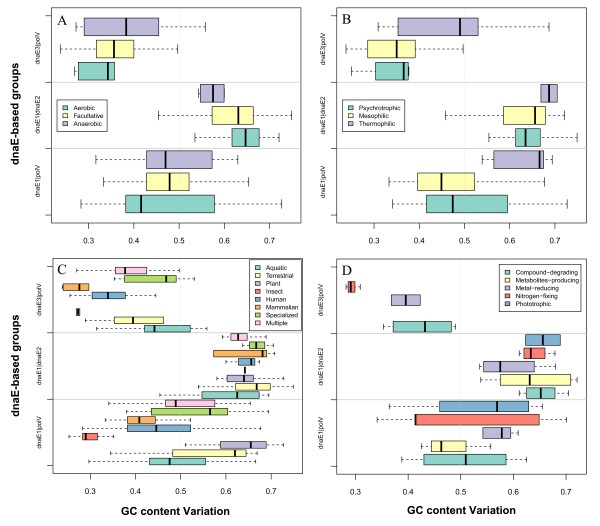
Mutator genes, especially those with dominant effects on the mutation spectra, are biased towards either GC or AT richness, and they alter genomic GC content in the two opposite directions. Increased bacterial genome size (or gene number) appears to rely on increased genomic GC content; however, it is unclear whether the changes are directly related to certain environmental pressures. Certain environmental and bacteriological features are related to GC content variation, but their trends are more obvious when analyzed under the dnaE-based grouping scheme. Most terrestrial, plant-associated, and nitrogen-fixing bacteria are members of the dnaE1|dnaE2 group, whereas most pathogenic or symbiotic bacteria in insects, and those dwelling in aquatic environments, are largely members of the dnaE1|polV group.
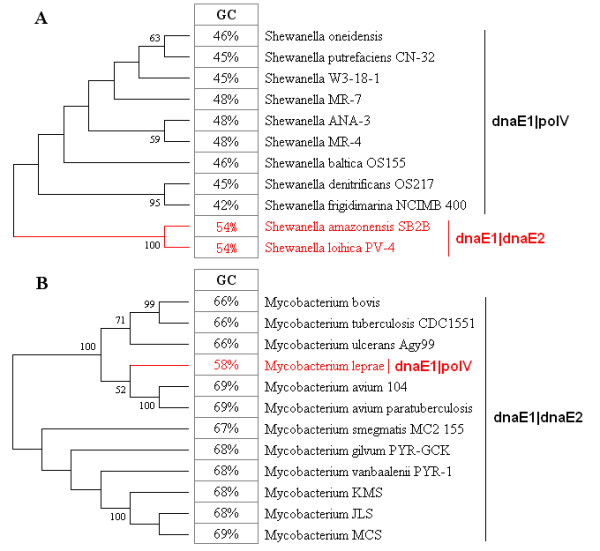
Our studies provide several lines of evidence indicating that DNA polymerase III α subunit and its isoforms participating in either replication (such as polC) or SOS mutagenesis/translesion synthesis (such as dnaE2), play dominant roles in determining GC variability. Other environmental or bacteriological factors, such as genome size, temperature, oxygen requirement, and habitat, either play subsidiary roles or rely indirectly on different mutator genes to fine-tune the GC content. These results provide a comprehensive insight into mechanisms of GC content variation and the robustness of eubacterial genomes in adapting their ever-changing environments over billions of years.
作为基因组序列变异的一个关键参数,细菌基因组的 GC 含量已经被研究了半个多世纪,提出了许多假说来解释这种 GC 含量的变化及其与其他基本过程的关系。以前,我们根据一个假说,将真细菌分为基于 dnaE 的组(DNA 聚合酶 IIIα亚基的二聚体组合),该假说认为 GC 含量的变化主要受基因组复制和 DNA 修复机制的控制。进一步的研究发现,两个主要的突变基因 polC 和 dnaE2 可能是导致基因组 GC 含量变化的原因。因此,我们进行了深入分析,以评估与真细菌基因组 GC 含量变化相关的各种潜在内在和外在因素。
突变基因,特别是那些对突变谱有显性影响的基因,偏向于 GC 或 AT 丰富度,它们以相反的两个方向改变基因组的 GC 含量。细菌基因组大小(或基因数量)的增加似乎依赖于基因组 GC 含量的增加;然而,尚不清楚这些变化是否与某些环境压力直接相关。某些环境和细菌学特征与 GC 含量的变化有关,但在基于 dnaE 的分组方案下进行分析时,它们的趋势更为明显。大多数陆地、植物相关和固氮细菌是 dnaE1|dnaE2 组的成员,而大多数昆虫中的致病性或共生细菌以及水生环境中的细菌,主要是 dnaE1|polV 组的成员。
我们的研究提供了几条证据表明,DNA 聚合酶 IIIα亚基及其参与复制(如 polC)或 SOS 诱变/跨损伤合成(如 dnaE2)的同工酶,在决定 GC 变异性方面起着主导作用。其他环境或细菌学因素,如基因组大小、温度、需氧性和栖息地,要么起着辅助作用,要么间接地依赖于不同的突变基因来微调 GC 含量。这些结果提供了对 GC 含量变化机制以及真细菌基因组在数十亿年中适应不断变化的环境的稳健性的全面了解。