Interfaculty Institute of Biochemistry, University of Tübingen, Tübingen, Germany.
PLoS Pathog. 2013;9(10):e1003688. doi: 10.1371/journal.ppat.1003688. Epub 2013 Oct 10.
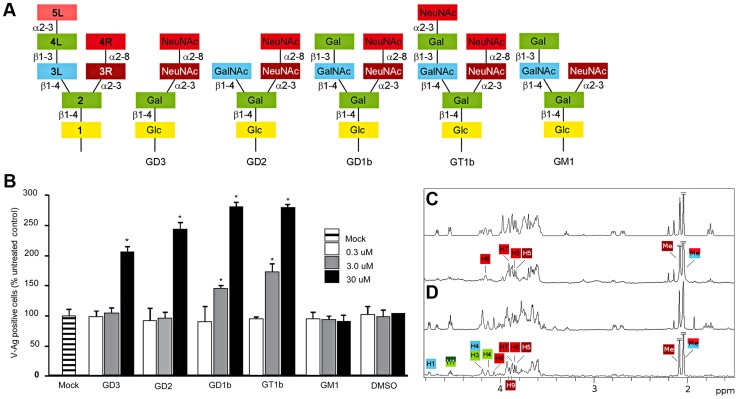
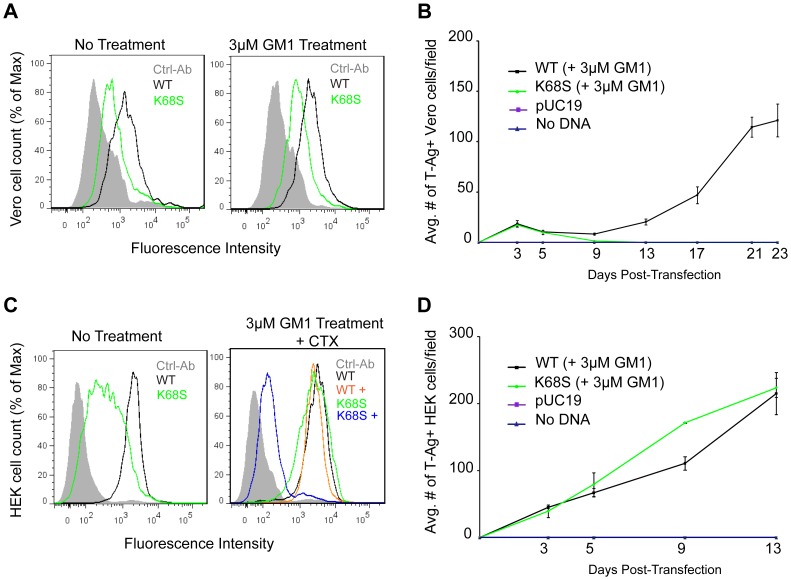
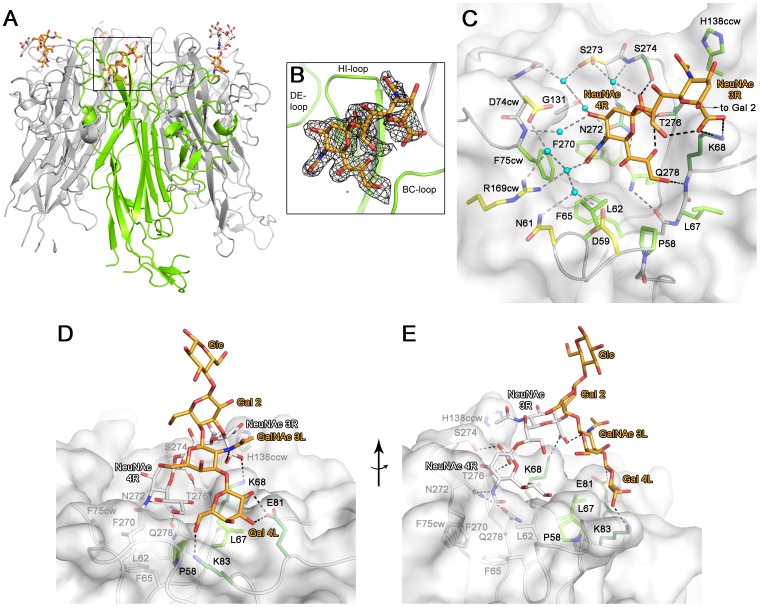
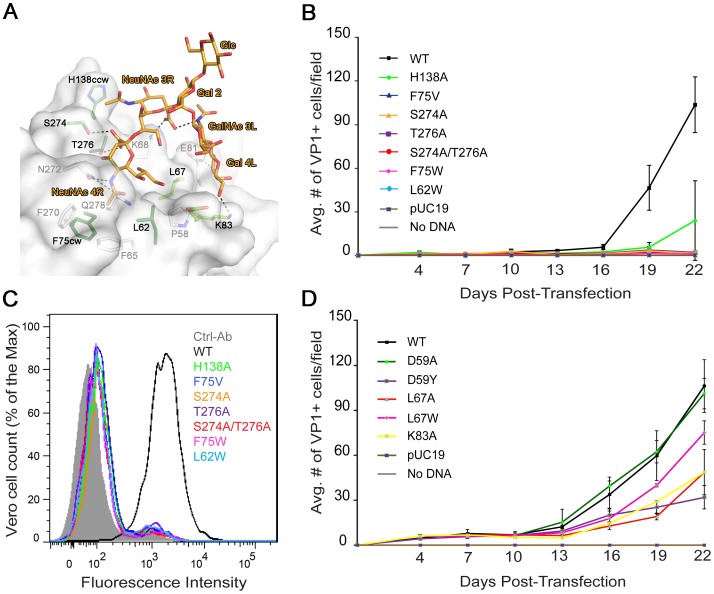
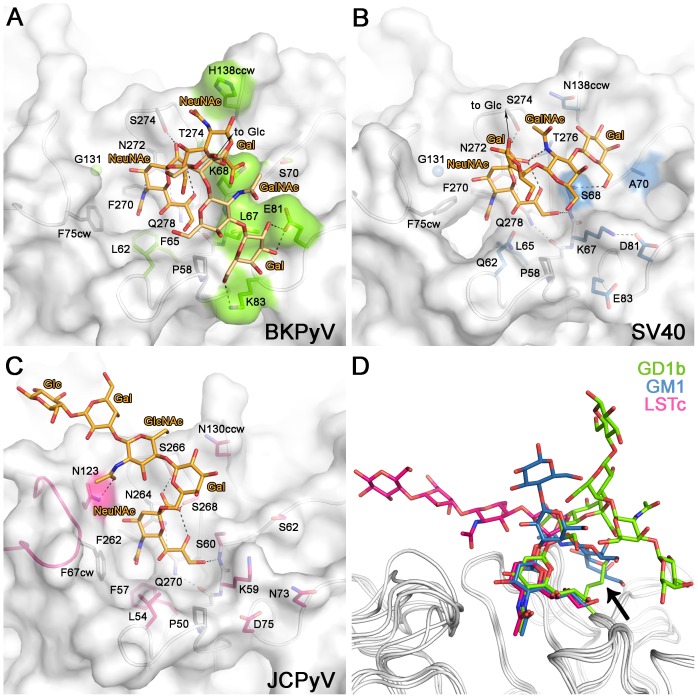
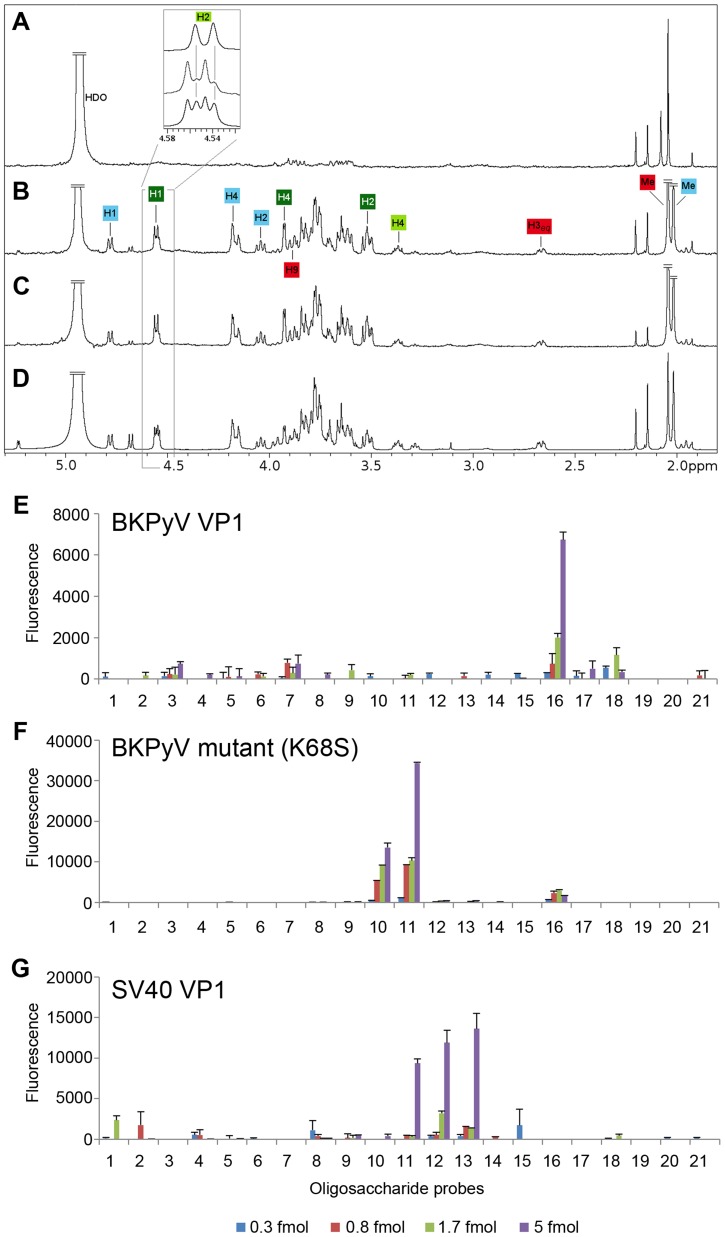
Viruses within a family often vary in their cellular tropism and pathogenicity. In many cases, these variations are due to viruses switching their specificity from one cell surface receptor to another. The structural requirements that underlie such receptor switching are not well understood especially for carbohydrate-binding viruses, as methods capable of structure-specificity studies are only relatively recently being developed for carbohydrates. We have characterized the receptor specificity, structure and infectivity of the human polyomavirus BKPyV, the causative agent of polyomavirus-associated nephropathy, and uncover a molecular switch for binding different carbohydrate receptors. We show that the b-series gangliosides GD3, GD2, GD1b and GT1b all can serve as receptors for BKPyV. The crystal structure of the BKPyV capsid protein VP1 in complex with GD3 reveals contacts with two sialic acid moieties in the receptor, providing a basis for the observed specificity. Comparison with the structure of simian virus 40 (SV40) VP1 bound to ganglioside GM1 identifies the amino acid at position 68 as a determinant of specificity. Mutation of this residue from lysine in BKPyV to serine in SV40 switches the receptor specificity of BKPyV from GD3 to GM1 both in vitro and in cell culture. Our findings highlight the plasticity of viral receptor binding sites and form a template to retarget viruses to different receptors and cell types.
家族内的病毒在细胞嗜性和致病性方面经常存在差异。在许多情况下,这些变化是由于病毒将其特异性从一种细胞表面受体切换到另一种受体。这种受体切换所基于的结构要求尚不清楚,特别是对于碳水化合物结合病毒,因为能够进行结构特异性研究的方法只是在最近才被开发出来用于碳水化合物。我们已经描述了人类多瘤病毒 BKPyV 的受体特异性、结构和感染力,BKPyV 是多瘤病毒相关性肾病的病原体,并揭示了一种用于结合不同碳水化合物受体的分子开关。我们表明,b 系列神经节苷脂 GD3、GD2、GD1b 和 GT1b 均可作为 BKPyV 的受体。BKPyV 衣壳蛋白 VP1 与 GD3 复合物的晶体结构揭示了与受体中两个唾液酸部分的接触,为观察到的特异性提供了基础。与结合神经节苷脂 GM1 的猿猴病毒 40(SV40)VP1 的结构进行比较,确定了位置 68 处的氨基酸是特异性的决定因素。将该残基从 BKPyV 中的赖氨酸突变为 SV40 中的丝氨酸,可将 BKPyV 的受体特异性从 GD3 切换到 GM1,无论是在体外还是在细胞培养中都是如此。我们的发现强调了病毒受体结合位点的可塑性,并为将病毒靶向不同受体和细胞类型提供了模板。