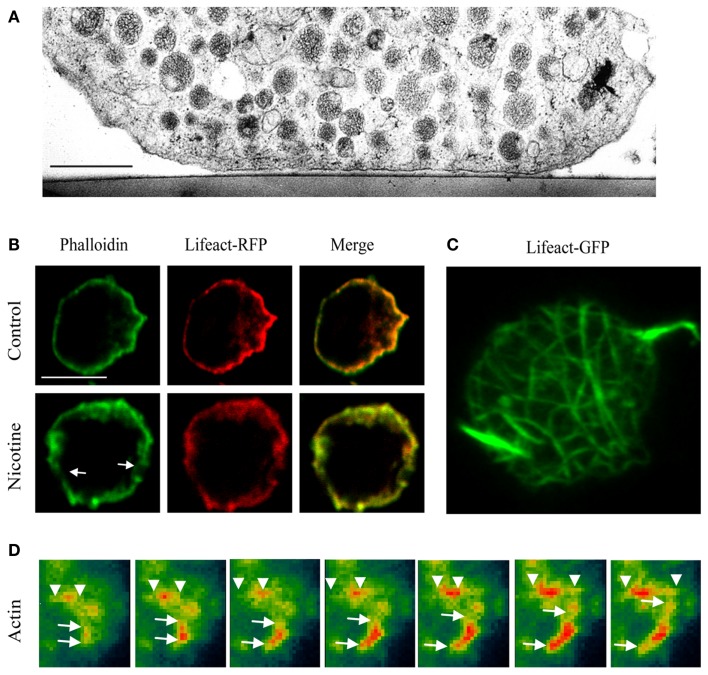
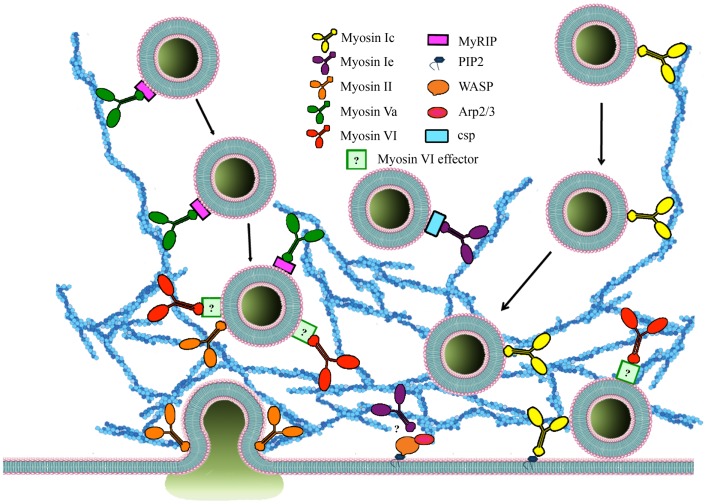
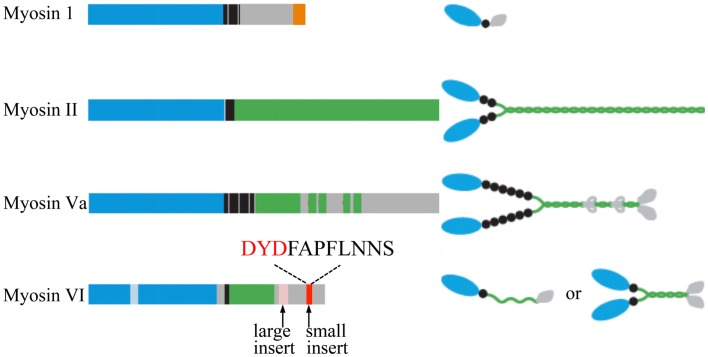
Papadopulos Andreas, Tomatis Vanesa M, Kasula Ravikiran, Meunier Frederic A
Queensland Brain Institute, The University of Queensland, St Lucia Campus , Brisbane, QLD , Australia.
Front Endocrinol (Lausanne). 2013 Oct 21;4:153. doi: 10.3389/fendo.2013.00153.
Dysregulation of regulated exocytosis is linked to an array of pathological conditions, including neurodegenerative disorders, asthma, and diabetes. Understanding the molecular mechanisms underpinning neuroexocytosis including the processes that allow neurosecretory vesicles to access and fuse with the plasma membrane and to recycle post-fusion, is therefore critical to the design of future therapeutic drugs that will efficiently tackle these diseases. Despite considerable efforts to determine the principles of vesicular fusion, the mechanisms controlling the approach of vesicles to the plasma membrane in order to undergo tethering, docking, priming, and fusion remain poorly understood. All these steps involve the cortical actin network, a dense mesh of actin filaments localized beneath the plasma membrane. Recent work overturned the long-held belief that the cortical actin network only plays a passive constraining role in neuroexocytosis functioning as a physical barrier that partly breaks down upon entry of Ca(2+) to allow secretory vesicles to reach the plasma membrane. A multitude of new roles for the cortical actin network in regulated exocytosis have now emerged and point to highly dynamic novel functions of key myosin molecular motors. Myosins are not only believed to help bring about dynamic changes in the actin cytoskeleton, tethering and guiding vesicles to their fusion sites, but they also regulate the size and duration of the fusion pore, thereby directly contributing to the release of neurotransmitters and hormones. Here we discuss the functions of the cortical actin network, myosins, and their effectors in controlling the processes that lead to tethering, directed transport, docking, and fusion of exocytotic vesicles in regulated exocytosis.
受调控的胞吐作用失调与一系列病理状况相关,包括神经退行性疾病、哮喘和糖尿病。因此,了解支撑神经胞吐作用的分子机制,包括神经分泌囊泡与质膜接触、融合以及融合后回收利用的过程,对于设计能够有效治疗这些疾病的未来治疗药物至关重要。尽管人们为确定囊泡融合的原理付出了巨大努力,但对于控制囊泡接近质膜以进行拴系、对接、引发和融合的机制仍知之甚少。所有这些步骤都涉及皮质肌动蛋白网络,这是位于质膜下方的密集肌动蛋白丝网格。最近的研究推翻了长期以来的观点,即皮质肌动蛋白网络在神经胞吐作用中仅起被动约束作用,作为一种物理屏障,在钙离子进入时部分分解,以允许分泌囊泡到达质膜。现在已经出现了皮质肌动蛋白网络在受调控胞吐作用中的许多新作用,并指向关键肌球蛋白分子马达的高度动态新功能。肌球蛋白不仅被认为有助于引起肌动蛋白细胞骨架的动态变化,将囊泡拴系并引导至其融合位点,而且还调节融合孔的大小和持续时间,从而直接促进神经递质和激素的释放。在这里,我们讨论皮质肌动蛋白网络、肌球蛋白及其效应器在控制受调控胞吐作用中导致胞吐囊泡拴系、定向运输、对接和融合的过程中的功能。