State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing, China ; University of Chinese Academy of Sciences, Beijing, China.
State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing, China.
PLoS One. 2014 Jan 8;9(1):e84842. doi: 10.1371/journal.pone.0084842. eCollection 2014.
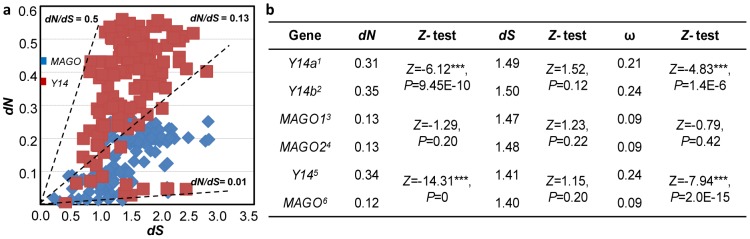
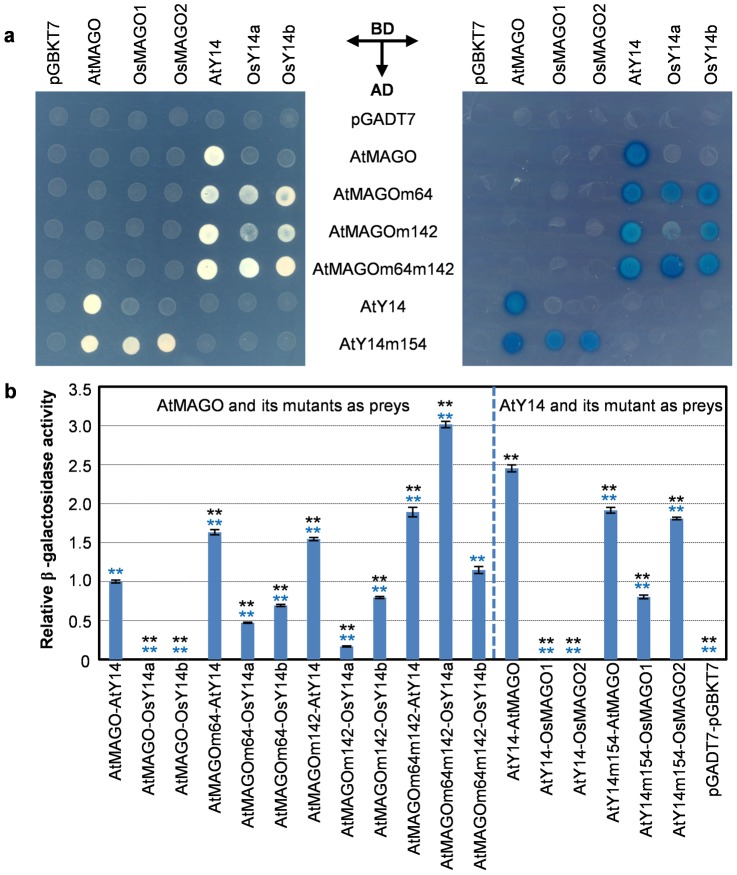
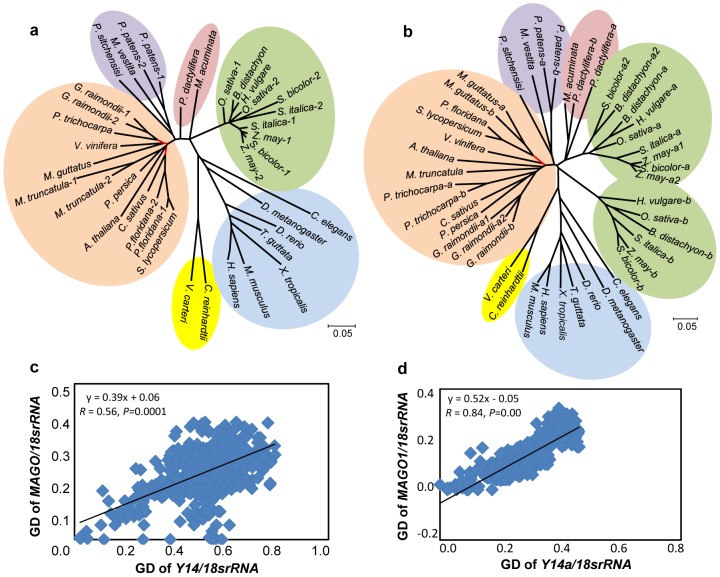
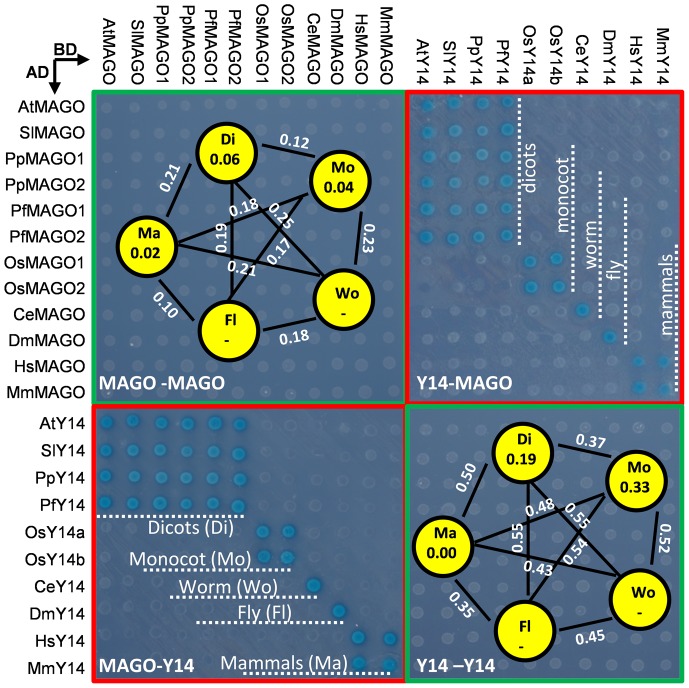
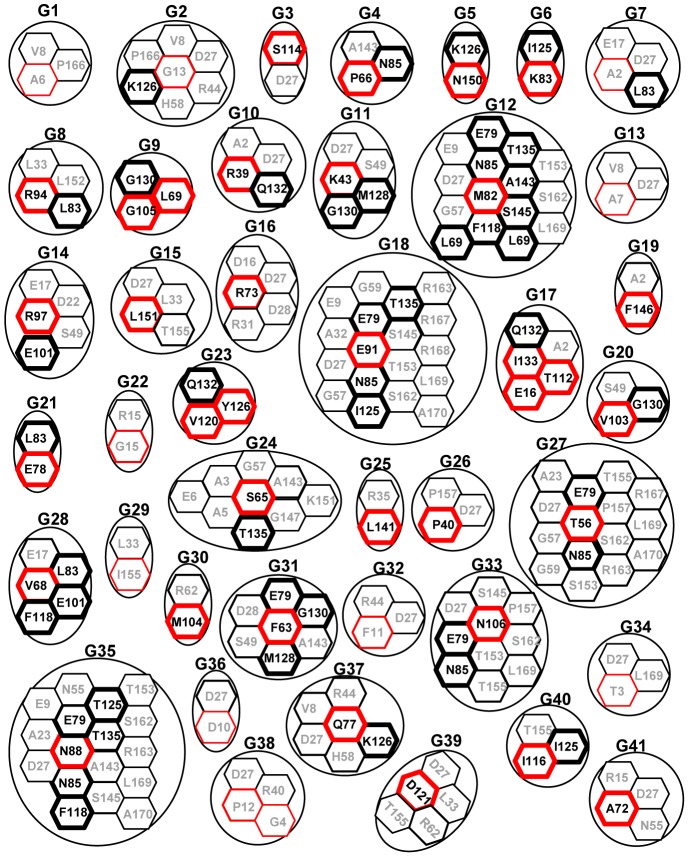
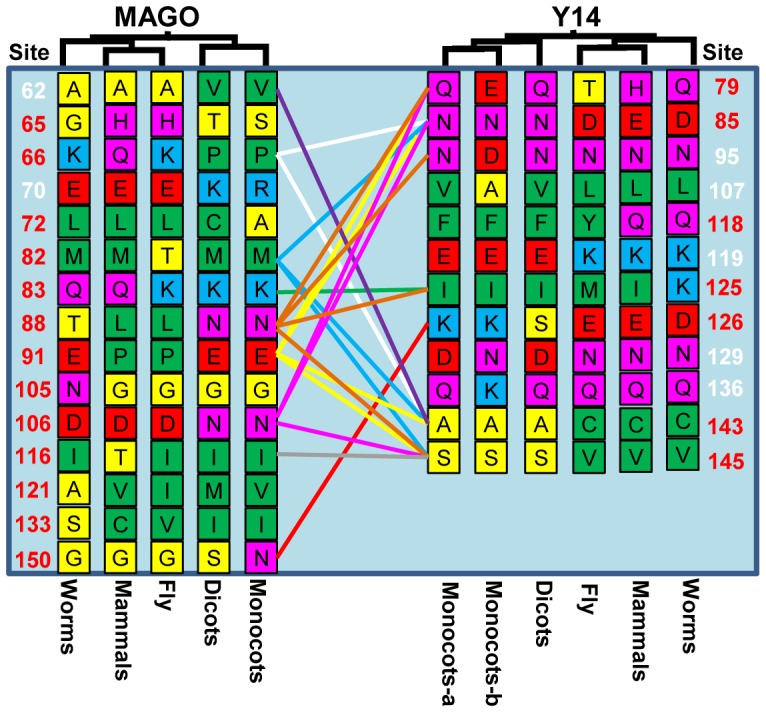
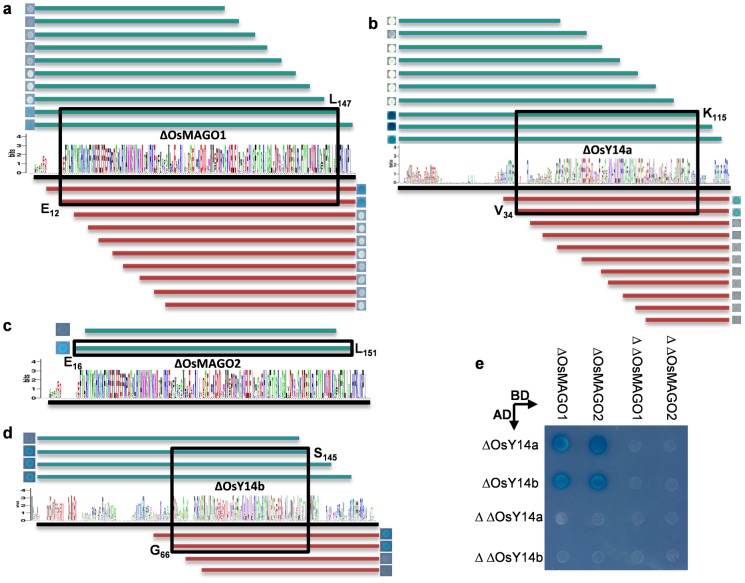
The exon junction complex (EJC) plays important roles in RNA metabolisms and the development of eukaryotic organisms. MAGO (short form of MAGO NASHI) and Y14 (also Tsunagi or RBM8) are the EJC core components. Their biological roles have been well investigated in various species, but the evolutionary patterns of the two gene families and their protein-protein interactions are poorly known. Genome-wide survey suggested that the MAGO and Y14 two gene families originated in eukaryotic organisms with the maintenance of a low copy. We found that the two protein families evolved slowly; however, the MAGO family under stringent purifying selection evolved more slowly than the Y14 family that was under relative relaxed purifying selection. MAGO and Y14 were obliged to form heterodimer in a eukaryotic organism, and this obligate mode was plesiomorphic. Lack of binding of MAGO to Y14 as functional barrier was observed only among distantly species, suggesting that a slow co-evolution of the two protein families. Inter-protein co-evolutionary signal was further quantified in analyses of the Tol-MirroTree and co-evolution analysis using protein sequences. About 20% of the 41 significantly correlated mutation groups (involving 97 residues) predicted between the two families was clade-specific. Moreover, around half of the predicted co-evolved groups and nearly all clade-specific residues fell into the minimal interaction domains of the two protein families. The mutagenesis effects of the clade-specific residues strengthened that the co-evolution is required for obligate MAGO-Y14 heterodimerization mode. In turn, the obliged heterodimerization in an organism serves as a strong functional constraint for the co-evolution of the MAGO and Y14 families. Such a co-evolution allows maintaining the interaction between the proteins through large evolutionary time scales. Our work shed a light on functional evolution of the EJC genes in eukaryotes, and facilitates to understand the co-evolutionary processes among protein families.
外显子衔接复合体(EJC)在 RNA 代谢和真核生物的发育中发挥重要作用。 MAGO(MAGO NASHI 的缩写)和 Y14(也称为 Tsunagi 或 RBM8)是 EJC 的核心成分。它们的生物学作用已在各种物种中得到很好的研究,但这两个基因家族的进化模式及其蛋白质-蛋白质相互作用知之甚少。全基因组调查表明,MAGO 和 Y14 两个基因家族起源于真核生物,其拷贝数维持在低水平。我们发现这两个蛋白质家族进化缓慢;然而,在严格的净化选择下,MAGO 家族的进化速度比相对宽松的净化选择下的 Y14 家族慢。 MAGO 和 Y14 必须在真核生物中形成异源二聚体,这种强制模式是原始的。只有在远缘物种中才观察到 MAGO 与 Y14 结合缺失作为功能障碍,这表明这两个蛋白质家族的进化速度较慢。在对 Tol-MirroTree 的蛋白质序列进行共进化分析和分析中,进一步量化了蛋白质间共进化的信号。在这两个家族之间预测的 41 个显著相关突变组(涉及 97 个残基)中,约有 20%是进化枝特异性的。此外,预测的共进化组中有近一半,几乎所有的进化枝特异性残基都落入了这两个蛋白质家族的最小相互作用结构域中。进化枝特异性残基的诱变效应增强了 MAGO-Y14 异源二聚体形成的共进化。反过来,在生物体中必需的异源二聚体化作为 MAGO 和 Y14 家族共进化的强大功能约束。这种共进化允许通过大的进化时间尺度来维持蛋白质之间的相互作用。我们的工作揭示了真核生物 EJC 基因的功能进化,并有助于理解蛋白质家族之间的共进化过程。