Laboratory of Plant Virology, Faculty of Agriculture, Saga University, Saga, Japan.
Laboratory of Plant Virology, Faculty of Agriculture, Saga University, Saga, Japan ; The United Graduate School of Agricultural Sciences, Kagoshima University, Kagoshima, Japan.
PLoS One. 2014 Jan 21;9(1):e85641. doi: 10.1371/journal.pone.0085641. eCollection 2014.
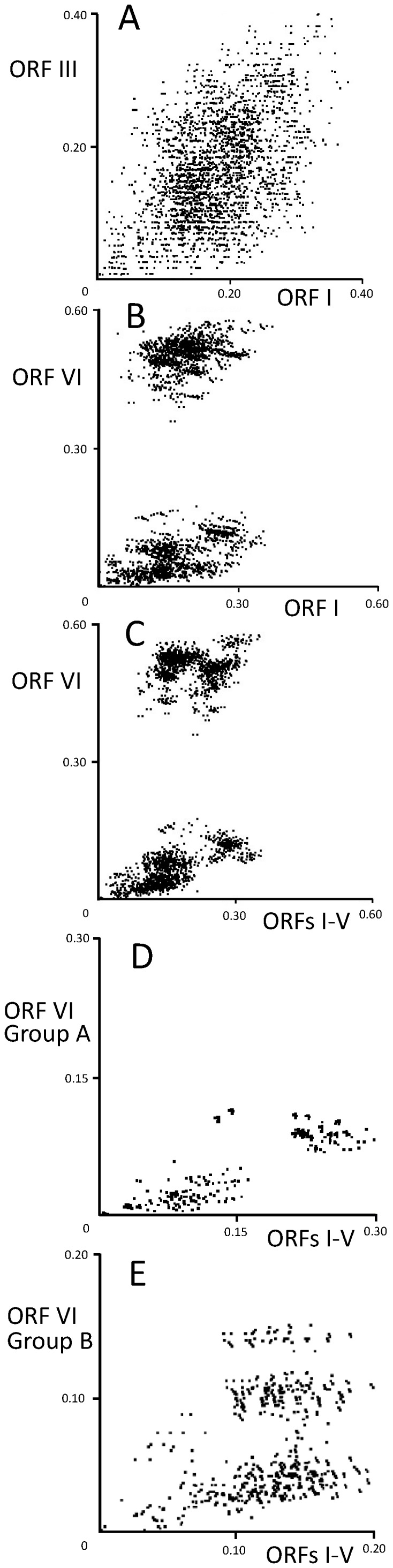
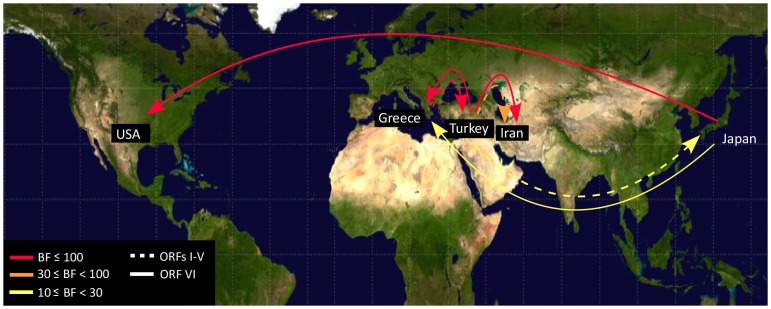
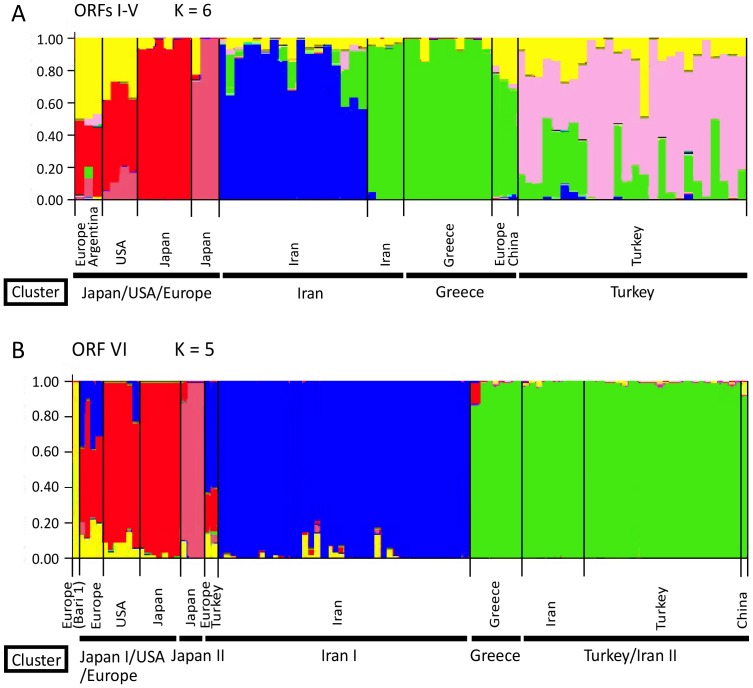
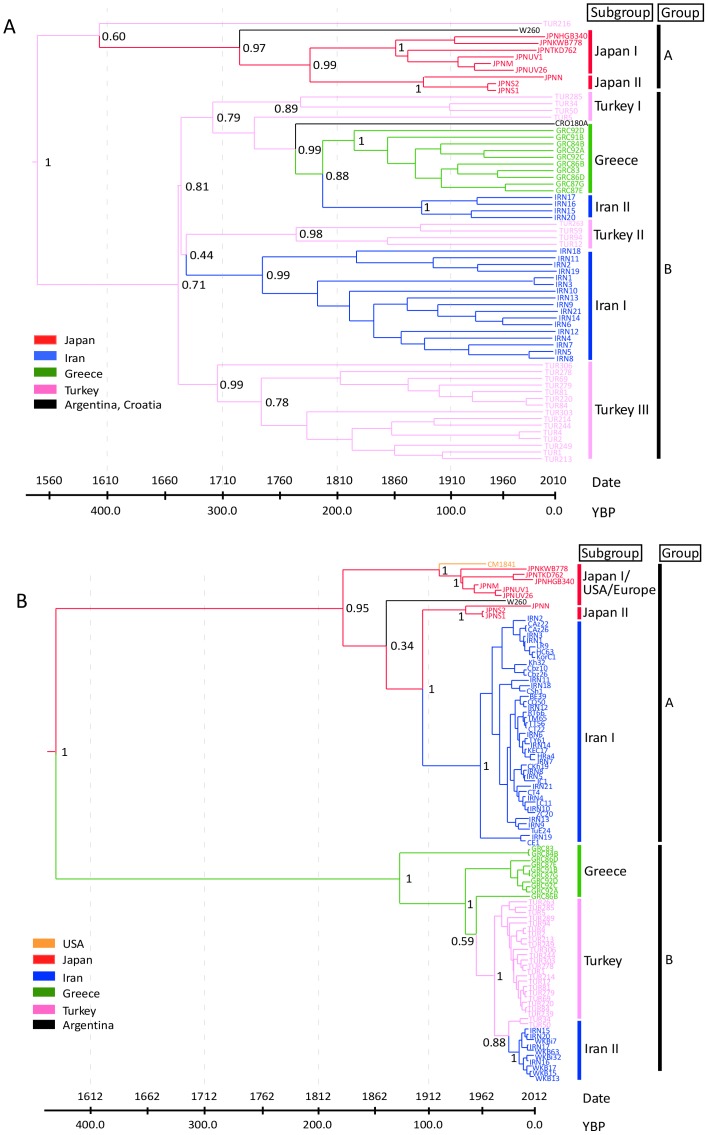
Cauliflower mosaic virus (CaMV) is a plant pararetrovirus with a double-stranded DNA genome. It is the type member of the genus Caulimovirus in the family Caulimoviridae. CaMV is transmitted by sap inoculation and in nature by aphids in a semi-persistent manner. To investigate the patterns and timescale of CaMV migration and evolution, we sequenced and analyzed the genomes of 67 isolates of CaMV collected mostly in Greece, Iran, Turkey, and Japan together with nine published sequences. We identified the open-reading frames (ORFs) in the genomes and inferred their phylogeny. After removing recombinant sequences, we estimated the substitution rates, divergence times, and phylogeographic patterns of the virus populations. We found that recombination has been a common feature of CaMV evolution, and that ORFs I-V have a different evolutionary history from ORF VI. The ORFs have evolved at rates between 1.71 and 5.81×10(-4) substitutions/site/year, similar to those of viruses with RNA or ssDNA genomes. We found four geographically confined lineages. CaMV probably spread from a single population to other parts of the world around 400-500 years ago, and is now widely distributed among Eurasian countries. Our results revealed evidence of frequent gene flow between populations in Turkey and those of its neighboring countries, with similar patterns observed for Japan and the USA. Our study represents the first report on the spatial and temporal spread of a plant pararetrovirus.
花椰菜花叶病毒 (CaMV) 是一种具有双链 DNA 基因组的植物拟反转录病毒。它是花椰菜花叶病毒科花椰菜花叶病毒属的模式成员。CaMV 通过汁液接种在植物间传播,在自然界中通过蚜虫以半持久方式传播。为了研究 CaMV 的迁移和进化模式和时间尺度,我们对主要在希腊、伊朗、土耳其和日本收集的 67 个 CaMV 分离株的基因组进行了测序和分析,同时还分析了 9 个已发表的序列。我们确定了基因组中的开放阅读框 (ORF),并推断了它们的系统发育。在去除重组序列后,我们估计了病毒群体的取代率、分歧时间和系统地理学模式。我们发现重组是 CaMV 进化的一个共同特征,并且 ORF I-V 与 ORF VI 具有不同的进化历史。ORF 以 1.71 到 5.81×10(-4)个替换/位点/年的速率进化,与具有 RNA 或 ssDNA 基因组的病毒相似。我们发现了四个具有地理局限性的谱系。CaMV 可能在 400-500 年前从一个单一种群传播到世界其他地区,现在在欧亚国家广泛分布。我们的研究结果揭示了土耳其及其邻国之间种群间频繁基因流动的证据,日本和美国也观察到了类似的模式。我们的研究代表了对植物拟反转录病毒时空传播的首次报道。