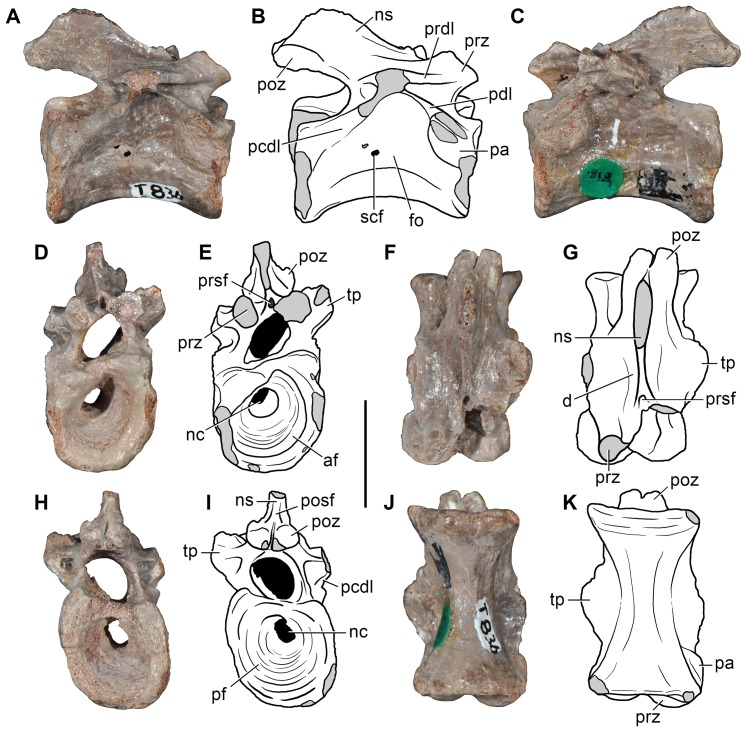
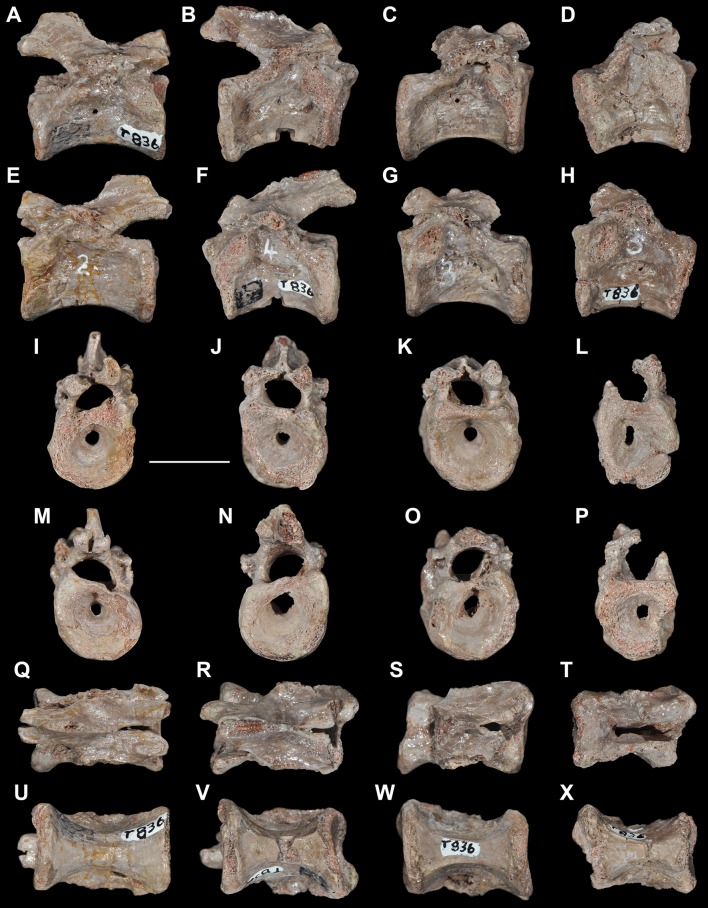
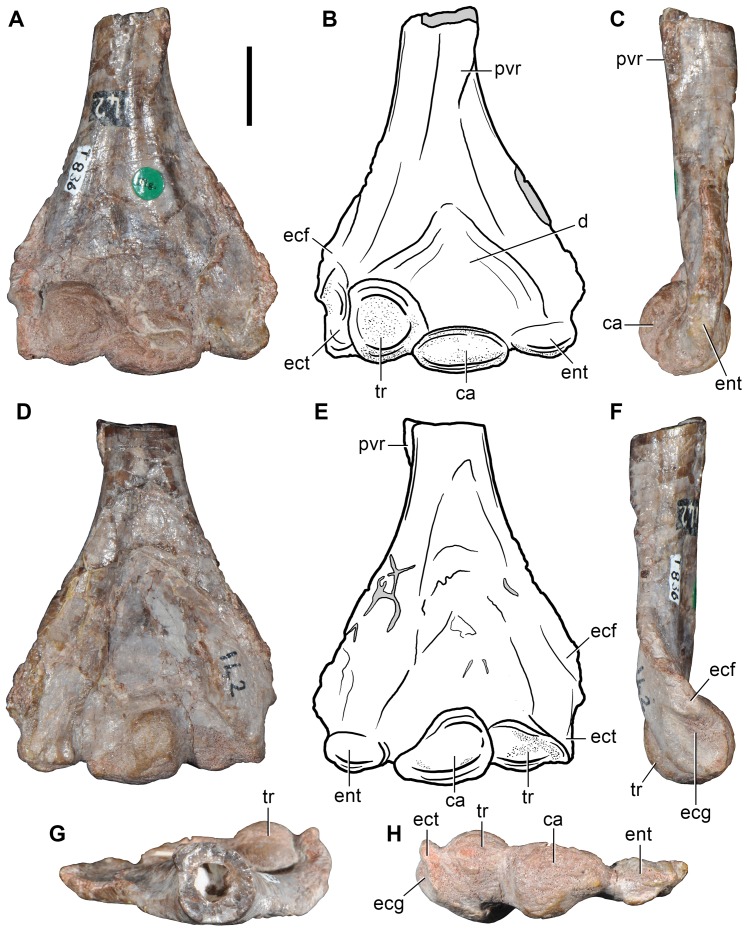
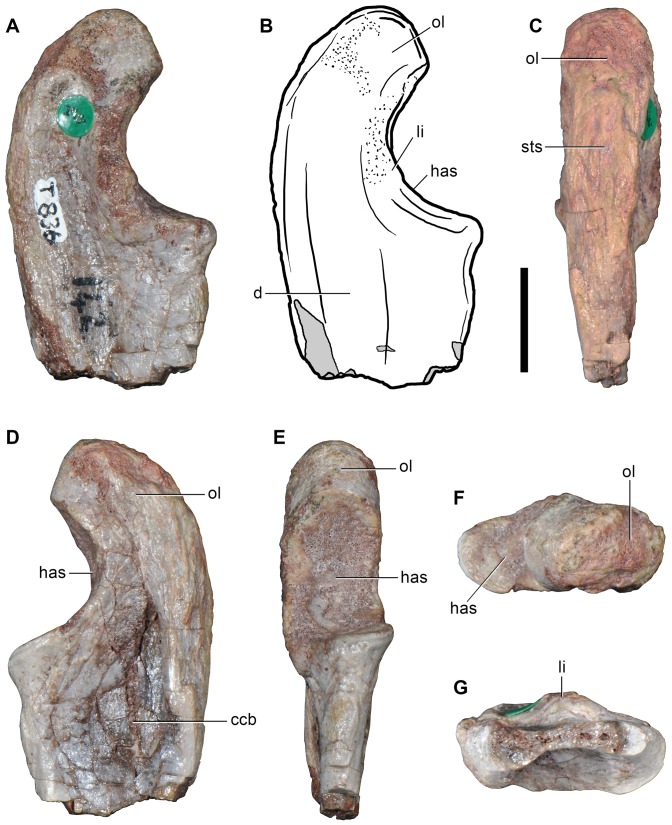
Ezcurra Martín D, Scheyer Torsten M, Butler Richard J
School of Geography, Earth and Environmental Sciences, University of Birmingham, Edgbaston, Birmingham, United Kingdom ; GeoBio-Center, Ludwig-Maximilian-Universität München, Munich, Germany.
Paläontologisches Institut und Museum, Universität Zürich, Zurich, Switzerland.
PLoS One. 2014 Feb 27;9(2):e89165. doi: 10.1371/journal.pone.0089165. eCollection 2014.
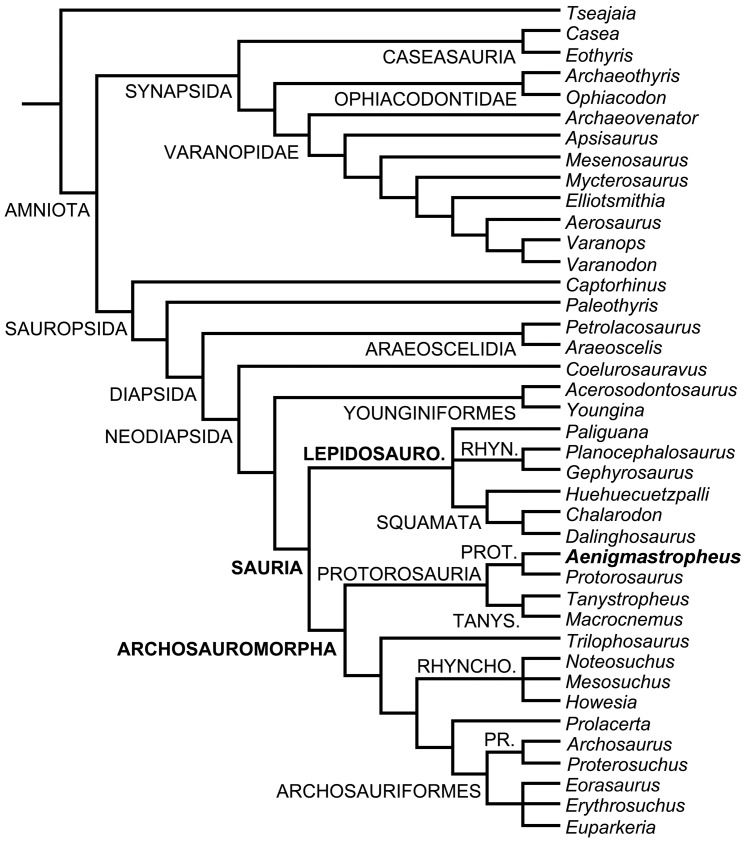
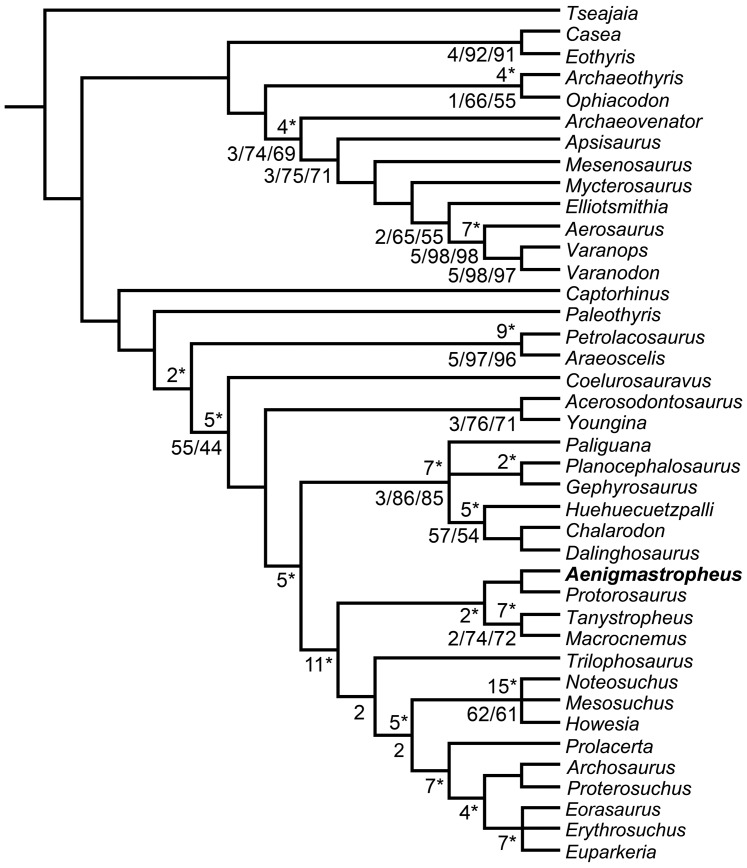
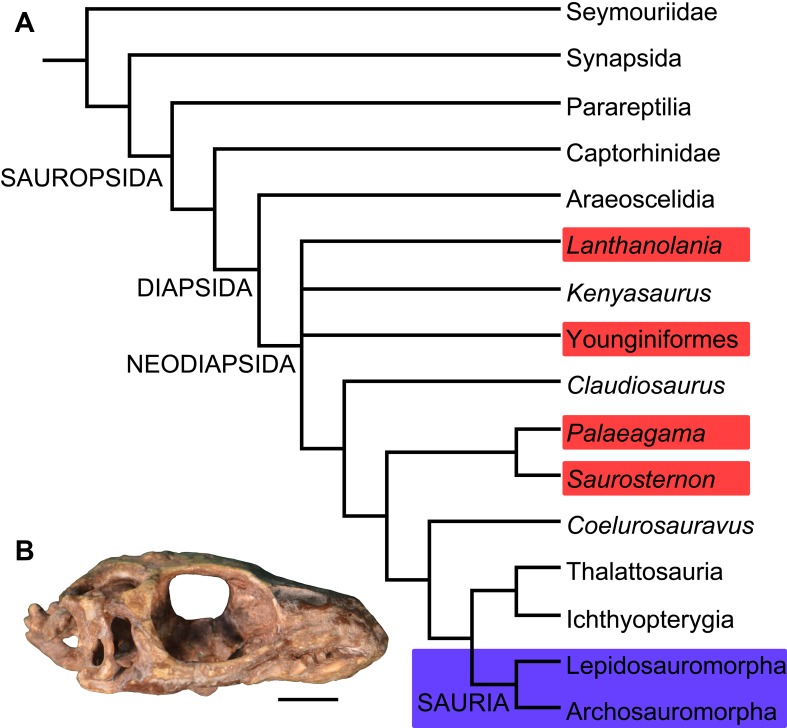
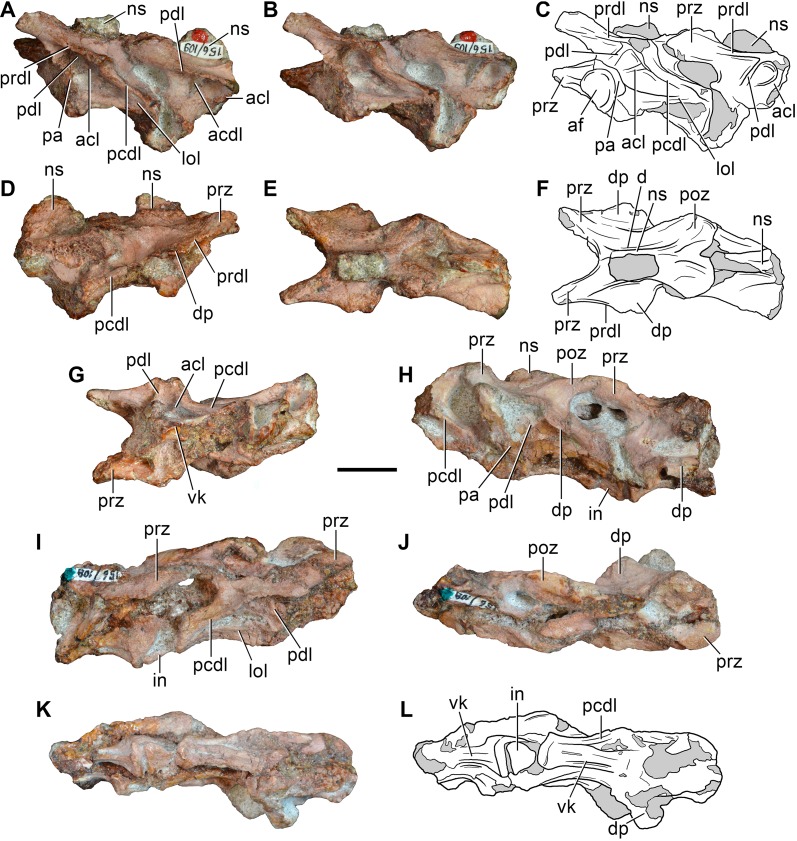
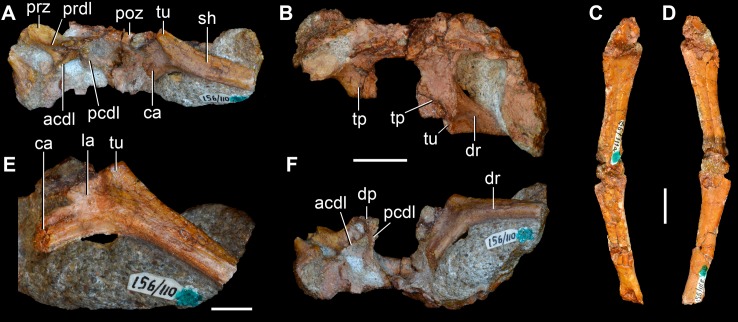
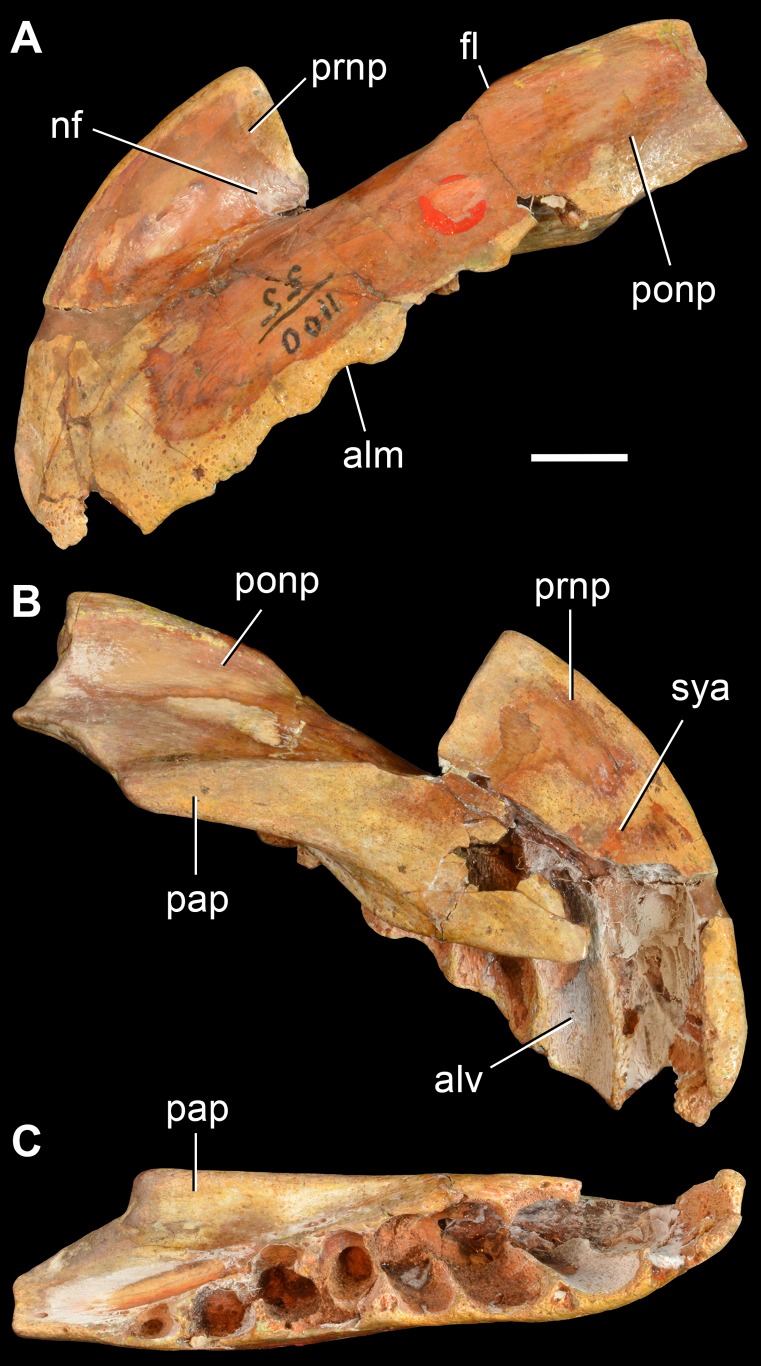
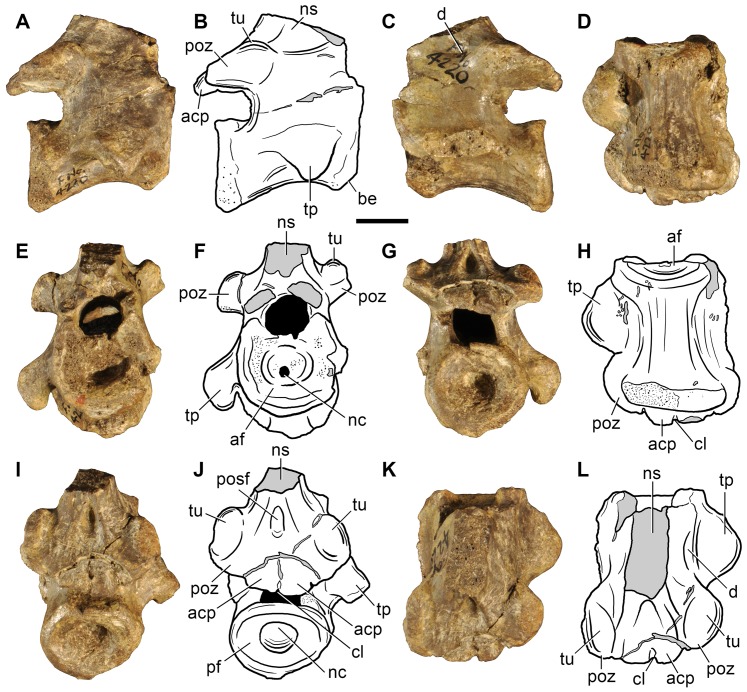
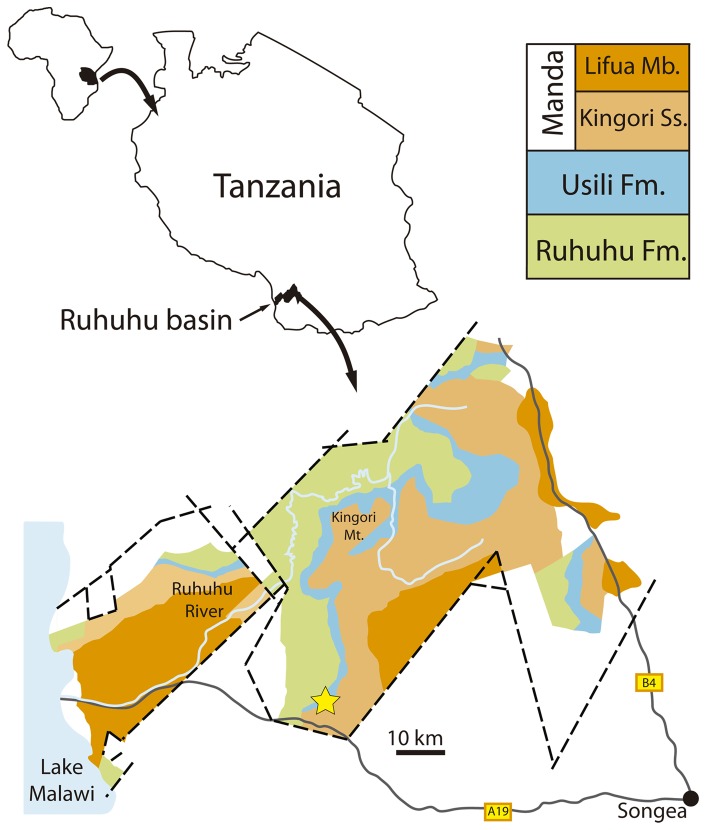
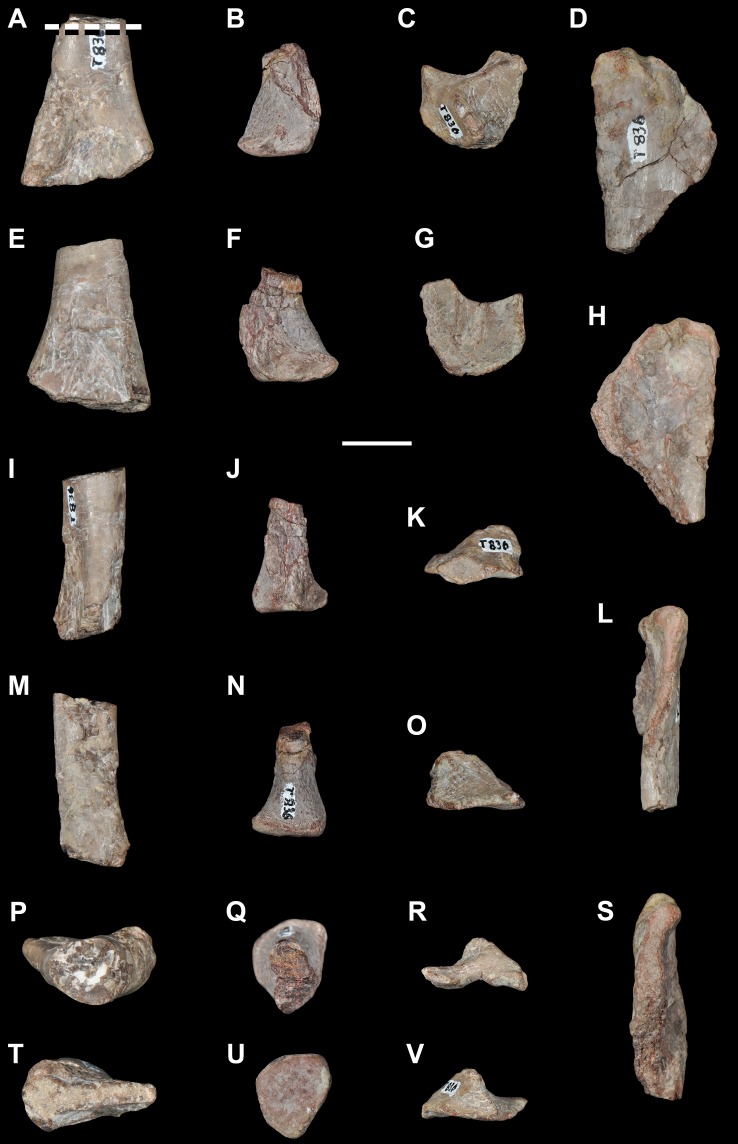
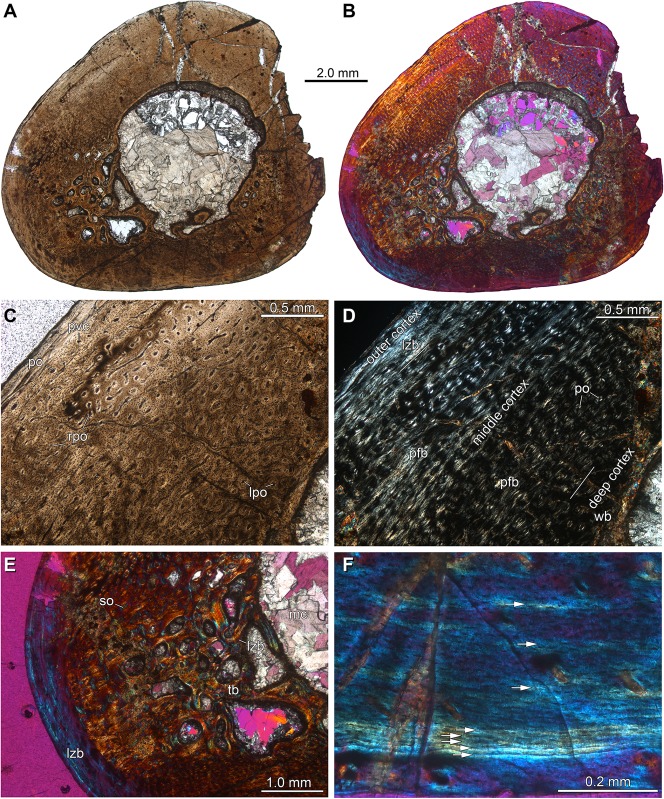
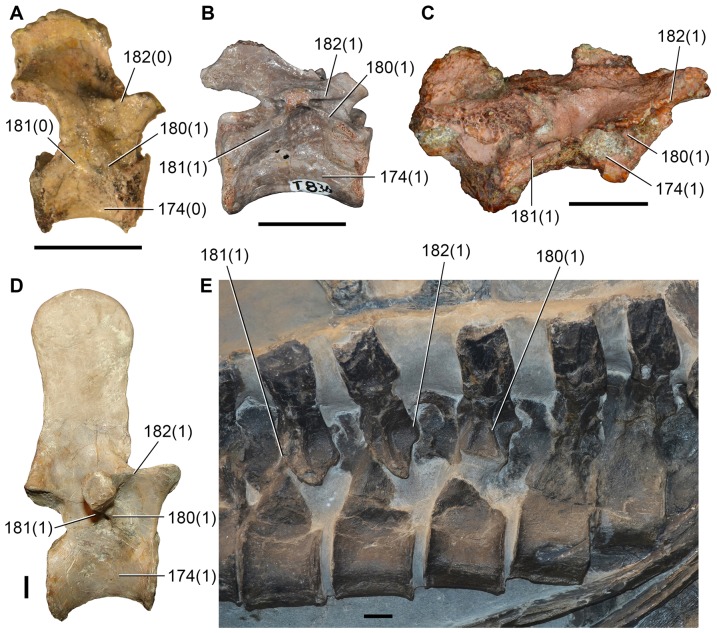
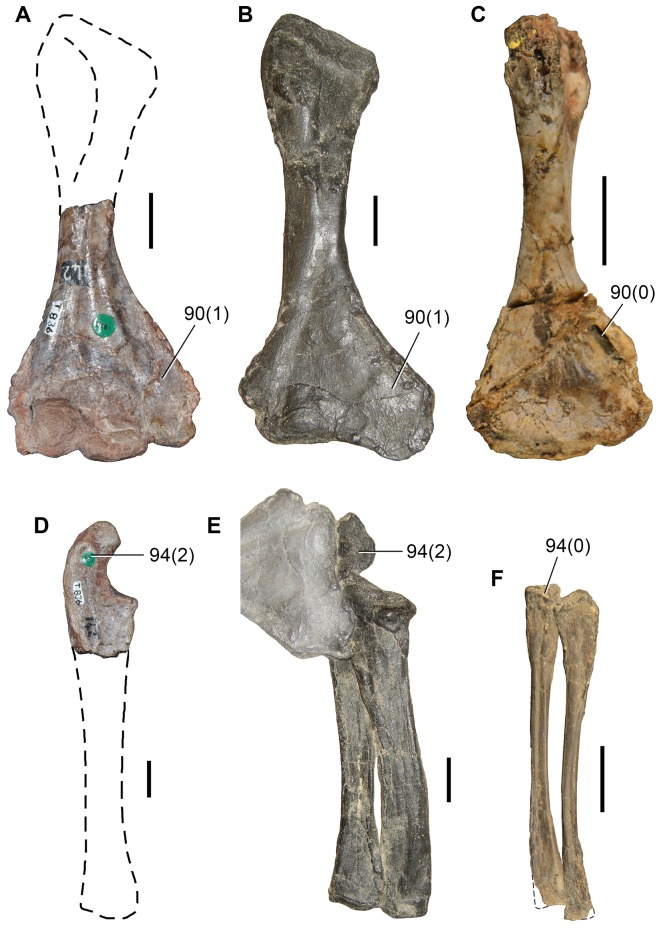
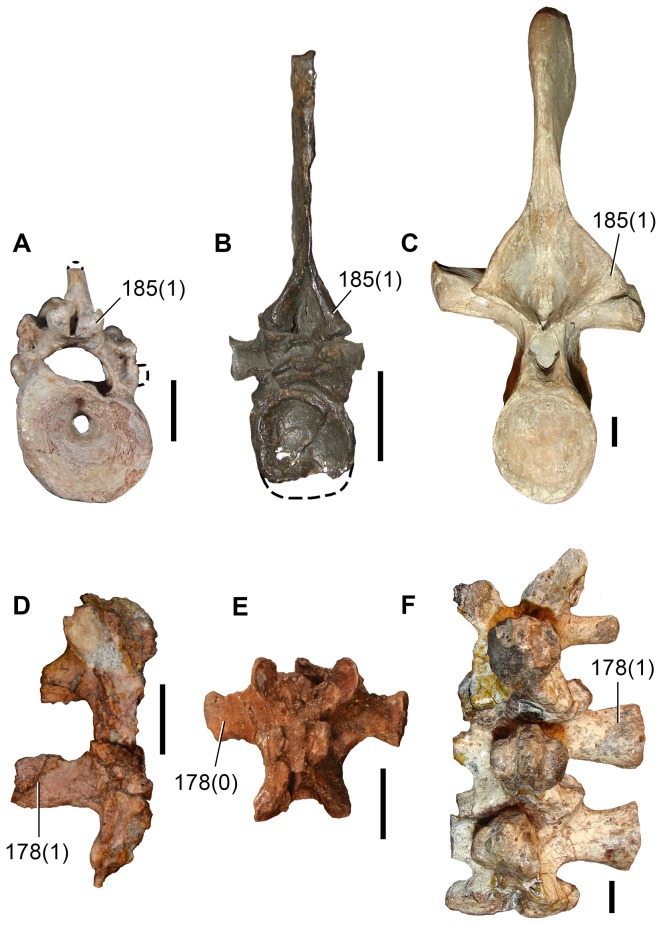
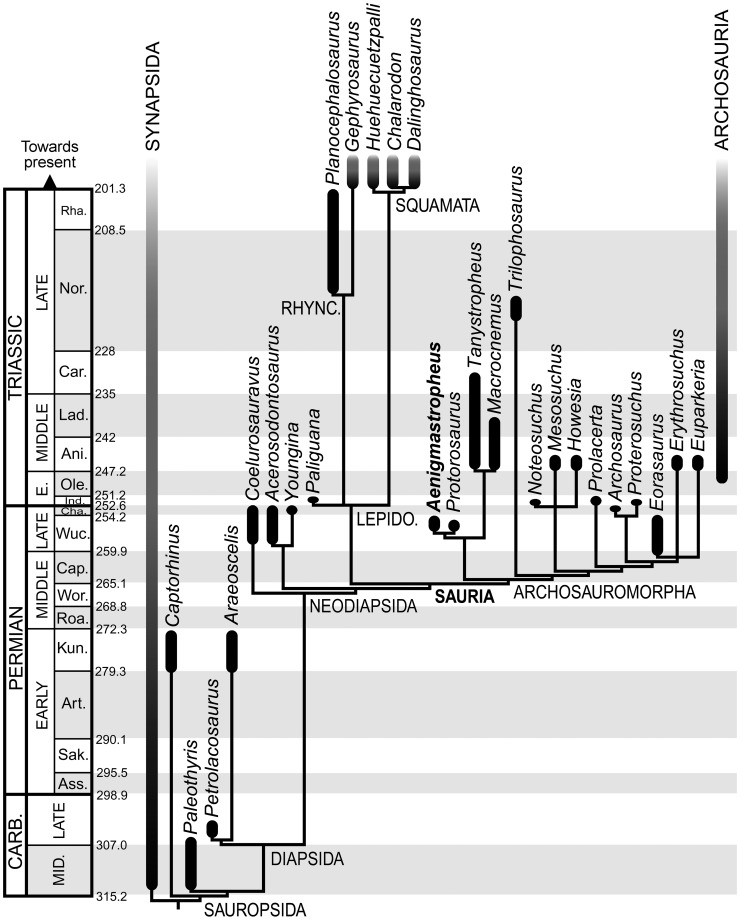
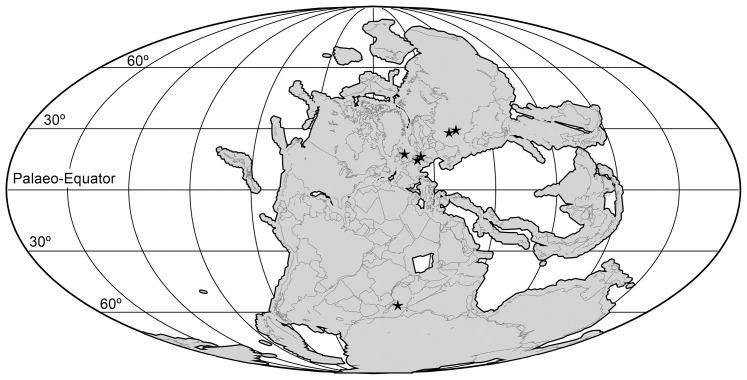
Sauria is the crown-group of Diapsida and is subdivided into Lepidosauromorpha and Archosauromorpha, comprising a high percentage of the diversity of living and fossil tetrapods. The split between lepidosauromorphs and archosauromorphs (the crocodile-lizard, or bird-lizard, divergence) is considered one of the key calibration points for molecular analyses of tetrapod phylogeny. Saurians have a very rich Mesozoic and Cenozoic fossil record, but their late Paleozoic (Permian) record is problematic. Several Permian specimens have been referred to Sauria, but the phylogenetic affinity of some of these records remains questionable. We reexamine and review all of these specimens here, providing new data on early saurian evolution including osteohistology, and present a new morphological phylogenetic dataset. We support previous studies that find that no valid Permian record for Lepidosauromorpha, and we also reject some of the previous referrals of Permian specimens to Archosauromorpha. The most informative Permian archosauromorph is Protorosaurus speneri from the middle Late Permian of Western Europe. A historically problematic specimen from the Late Permian of Tanzania is redescribed and reidentified as a new genus and species of basal archosauromorph: Aenigmastropheus parringtoni. The supposed protorosaur Eorasaurus olsoni from the Late Permian of Russia is recovered among Archosauriformes and may be the oldest known member of the group but the phylogenetic support for this position is low. The assignment of Archosaurus rossicus from the latest Permian of Russia to the archosauromorph clade Proterosuchidae is supported. Our revision suggests a minimum fossil calibration date for the crocodile-lizard split of 254.7 Ma. The occurrences of basal archosauromorphs in the northern (30°N) and southern (55°S) parts of Pangea imply a wider paleobiogeographic distribution for the group during the Late Permian than previously appreciated. Early archosauromorph growth strategies appear to be more diverse than previously suggested based on new data on the osteohistology of Aenigmastropheus.
蜥蜴类是双孔亚纲的冠群,可细分为鳞龙形下纲和主龙形下纲,在现存和化石四足动物的多样性中占比很高。鳞龙形下纲和主龙形下纲之间的分化(即鳄蜥或鸟蜥的分化)被认为是四足动物系统发育分子分析的关键校准点之一。蜥蜴类有非常丰富的中生代和新生代化石记录,但它们晚古生代(二叠纪)的记录存在问题。有几个二叠纪标本被归入蜥蜴类,但其中一些记录的系统发育亲缘关系仍存疑问。我们在此重新审视和回顾所有这些标本,提供包括骨组织学在内的早期蜥蜴类演化的新数据,并提出一个新的形态学系统发育数据集。我们支持先前的研究结果,即没有有效的二叠纪鳞龙形下纲记录,并且我们也拒绝了一些先前将二叠纪标本归入主龙形下纲的归类。最具信息价值的二叠纪主龙形下纲动物是来自西欧晚二叠世中期的斯氏原龙。对一个来自坦桑尼亚晚二叠世的历史上有问题的标本进行了重新描述,并重新鉴定为一种基础主龙形下纲的新属新种:帕林顿神秘鳄。来自俄罗斯晚二叠世的所谓原龙类奥尔森始龙被归入主龙形类,可能是该类群已知最古老的成员,但这一位置的系统发育支持度较低。来自俄罗斯最新二叠世的罗斯西鳄归入主龙形下纲原鳄科的归类得到支持。我们的修订表明,鳄蜥分化的最小化石校准日期为2.547亿年前。泛大陆北部(北纬30°)和南部(南纬55°)出现基础主龙形下纲动物,这意味着该类群在晚二叠世的古生物地理分布比之前认为的更广。根据神秘鳄骨组织学的新数据,早期主龙形下纲的生长策略似乎比之前认为的更加多样。