Department of Medicine (Microbiology and Immunobiology), Division of Infectious Diseases, Massachusetts General Hospital and Harvard Medical School, Cambridge, Massachusetts, United States of America.
PLoS One. 2014 Apr 9;9(4):e93461. doi: 10.1371/journal.pone.0093461. eCollection 2014.
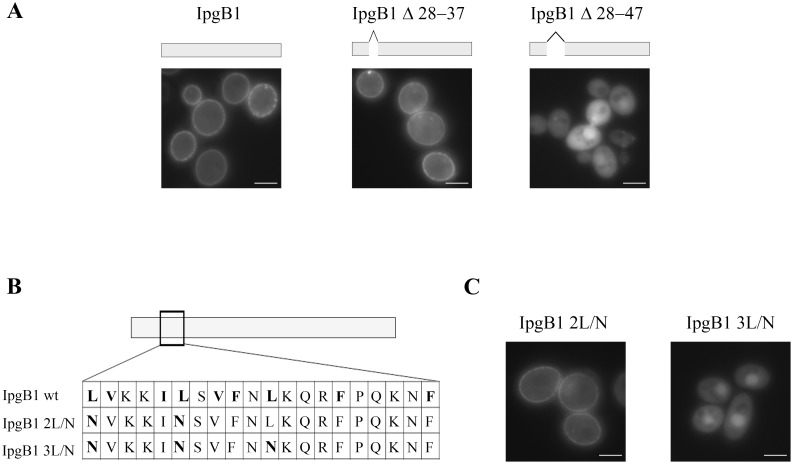
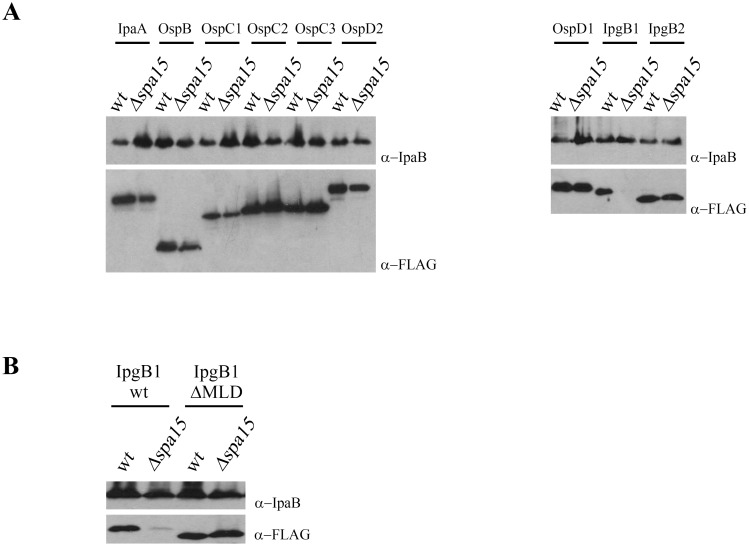
Type 3 secretion systems are complex nanomachines used by many Gram-negative bacteria to deliver tens of proteins (effectors) directly into host cells. Once delivered into host cells, effectors often target to specific cellular loci where they usurp host cell processes to their advantage. Here, using the yeast model system, we identify the membrane localization domain (MLD) of IpgB1, a stretch of 20 amino acids enriched for hydrophobic residues essential for the targeting of this effector to the plasma membrane. Embedded within these residues are ten that define the IpgB1 chaperone-binding domain for Spa15. As observed with dedicated class IA chaperones that mask hydrophobic MLDs, Spa15, a class IB chaperone, promotes IpgB1 stability by binding this hydrophobic region. However, despite being stable, an IpgB1 allele that lacks the MLD is not recognized as a secreted substrate. Similarly, deletion of the chaperone binding domains of IpgB1 and three additional Spa15-dependent effectors result in alleles that are no longer recognized as secreted substrates despite the presence of intact N-terminal secretion signal sequences. This is in contrast with MLD-containing effectors that bind class IA dedicated chaperones, as deletion of the MLD of these effectors alleviates the chaperone requirement for secretion. These observations indicate that at least for substrates of class IB chaperones, the chaperone-effector complex plays a major role in defining type 3 secreted proteins and highlight how a single region of an effector can play important roles both within prokaryotic and eukaryotic cells.
III 型分泌系统是一种复杂的纳米机器,许多革兰氏阴性菌利用它将数十种蛋白质(效应蛋白)直接输送到宿主细胞中。一旦进入宿主细胞,效应蛋白通常会靶向特定的细胞位置,在那里它们篡夺宿主细胞的过程以获取优势。在这里,我们使用酵母模型系统,确定了 IpgB1 的膜定位结构域(MLD),这是一段富含疏水性残基的 20 个氨基酸序列,对于将这种效应蛋白靶向质膜是必不可少的。在这些残基中,有十个残基定义了 IpgB1 的 Spa15 伴侣结合域。正如专门的 IA 类伴侣蛋白掩盖疏水性 MLD 所观察到的那样,Spa15 作为 IB 类伴侣蛋白,通过结合这个疏水区促进 IpgB1 的稳定性。然而,尽管 IpgB1 等位基因稳定,但缺乏 MLD 的等位基因不会被识别为分泌底物。同样,缺失 IpgB1 和三个额外的 Spa15 依赖性效应物的伴侣结合域会导致等位基因不再被识别为分泌底物,尽管存在完整的 N 端分泌信号序列。这与含有 MLD 的效应物形成对比,这些效应物结合 IA 类专门的伴侣蛋白,因为这些效应物的 MLD 缺失减轻了伴侣蛋白对分泌的需求。这些观察结果表明,至少对于 IB 类伴侣蛋白的底物,伴侣-效应物复合物在定义 III 型分泌蛋白方面起着重要作用,并强调了效应物的一个单一区域如何在原核和真核细胞中发挥重要作用。