Department of Pharmacology and Molecular Sciences, The Johns Hopkins University School of Medicine, Baltimore, MD 21205.
Department of Chemistry, Rutgers University, Newark, New Jersey 07102.
FEBS J. 2014 Jun;281(12):2820-2837. doi: 10.1111/febs.12823. Epub 2014 May 12.
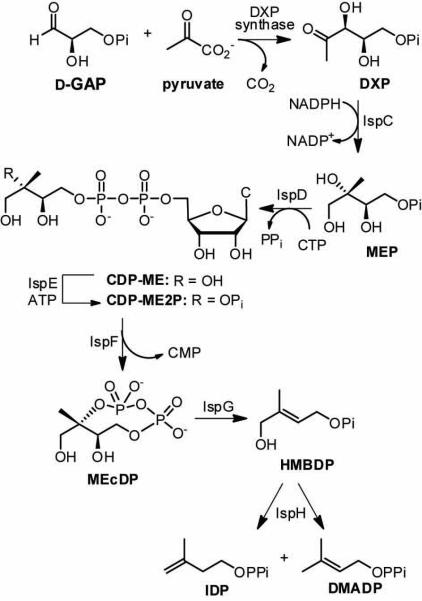
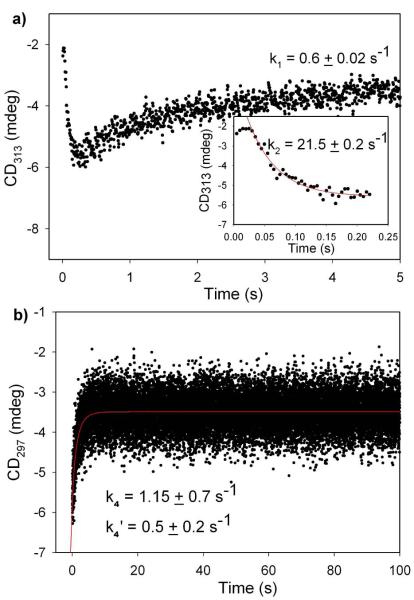
1-Deoxy-D-xylulose 5-phosphate (DXP) synthase catalyzes the formation of DXP from pyruvate and D-glyceraldehyde 3-phosphate (GraP) in a thiamin diphosphate-dependent manner, and is the first step in the essential pathway to isoprenoids in human pathogens. Understanding the mechanism of this unique enzyme is critical for developing new anti-infective agents that selectively target isoprenoid biosynthesis. The present study used mutagenesis and a combination of protein fluorescence, CD and kinetics experiments to investigate the roles of Arg420, Arg478 and Tyr392 in substrate binding and catalysis. The results support a random sequential, preferred order mechanism, and predict that Arg420 and Arg478 are involved in binding of the acceptor substrate, GraP. D-Glyceraldehyde, an alternative acceptor substrate lacking the phosphoryl group predicted to interact with Arg420 and Arg478, also accelerates decarboxylation of the predecarboxylation intermediate C2α-lactylthiamin diphosphate (LThDP) on DXP synthase, indicating that this binding interaction is not absolutely required, and that the hydroxyaldehyde sufficiently triggers decarboxylation. Unexpectedly, Tyr392 contributes to GraP affinity, and is not required for LThDP formation or its GraP-promoted decarboxylation. Time-resolved CD spectroscopy and NMR experiments indicate that LThDP is significantly stabilized on R420A and Y392F variants as compared with wild-type DXP synthase in the absence of acceptor substrate, but these substitutions do not appear to affect the rate of GraP-promoted LThDP decarboxylation in the presence of high levels of GraP, and LThDP formation remains the rate-limiting step. These results suggest a role of these residues in promoting GraP binding, which in turn facilitates decarboxylation, and also highlight interesting differences between DXP synthase and other thiamin diphosphate-dependent enzymes.
1-1-脱氧-D-木酮糖 5-磷酸(DXP)合酶以硫胺素二磷酸依赖性的方式催化丙酮酸和 D-甘油醛 3-磷酸(GraP)生成 DXP,是人体病原体中类异戊二烯基本途径的第一步。了解这种独特酶的机制对于开发选择性靶向类异戊二烯生物合成的新型抗感染药物至关重要。本研究使用突变和蛋白质荧光、CD 和动力学实验的组合来研究 Arg420、Arg478 和 Tyr392 在底物结合和催化中的作用。结果支持随机顺序、优先顺序机制,并预测 Arg420 和 Arg478 参与结合受体底物 GraP。D-甘油醛是一种替代的受体底物,缺乏预测与 Arg420 和 Arg478 相互作用的磷酸基团,也能加速 DXP 合酶上 C2α-乳酰硫胺素二磷酸(LThDP)的脱羧反应,表明这种结合相互作用不是绝对必需的,羟醛足以引发脱羧反应。出乎意料的是,Tyr392 有助于 GraP 的亲和力,并且不需要 LThDP 的形成或其在 GraP 促进下的脱羧反应。时间分辨 CD 光谱和 NMR 实验表明,与野生型 DXP 合酶相比,在没有受体底物的情况下,LThDP 在 R420A 和 Y392F 变体上显著稳定,但这些取代似乎并不影响在高浓度 GraP 存在下 GraP 促进的 LThDP 脱羧反应的速率,并且 LThDP 的形成仍然是限速步骤。这些结果表明这些残基在促进 GraP 结合中起作用,这反过来又促进脱羧反应,并且还突出了 DXP 合酶与其他硫胺素二磷酸依赖性酶之间的有趣差异。