Department of Botany II, Cologne Biocenter, University of Cologne Cologne, Germany ; Lophius Biosciences Regensburg, Germany.
Department of Botany II, Cologne Biocenter, University of Cologne Cologne, Germany ; Quintiles GmbH Neu-Isenburg, Germany.
Front Plant Sci. 2014 Apr 8;5:126. doi: 10.3389/fpls.2014.00126. eCollection 2014.
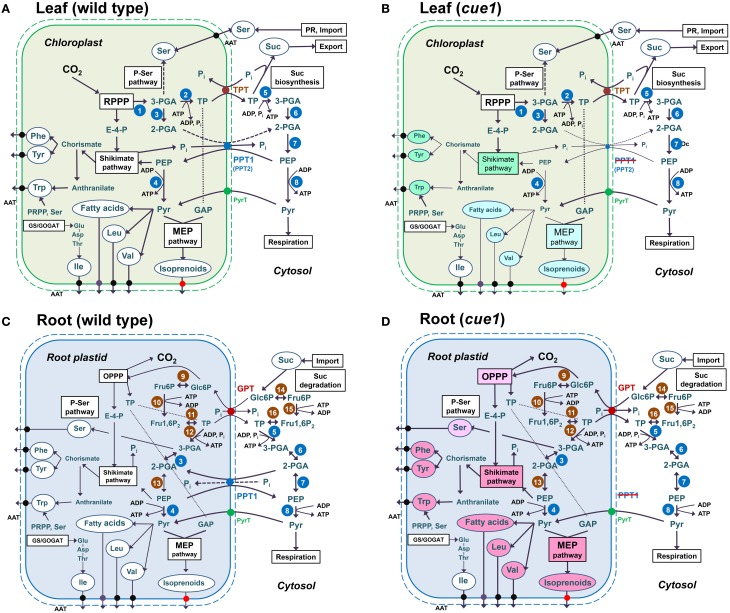
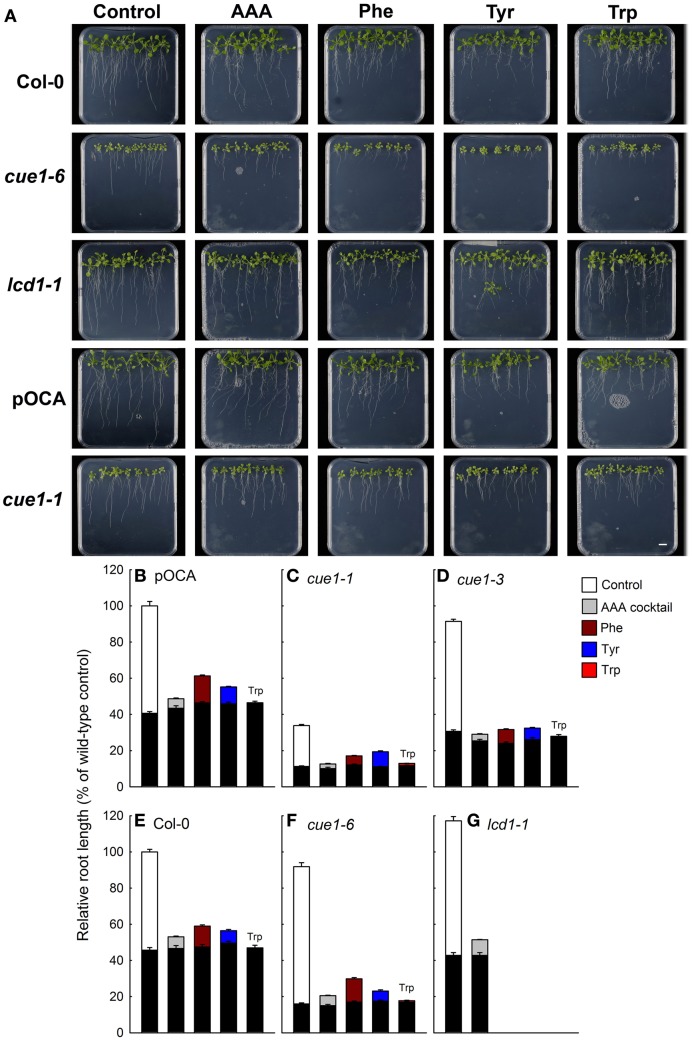
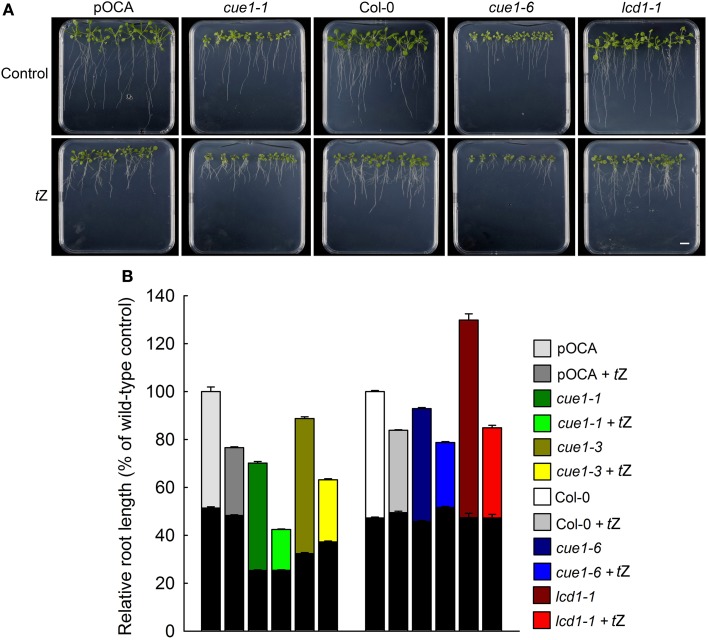
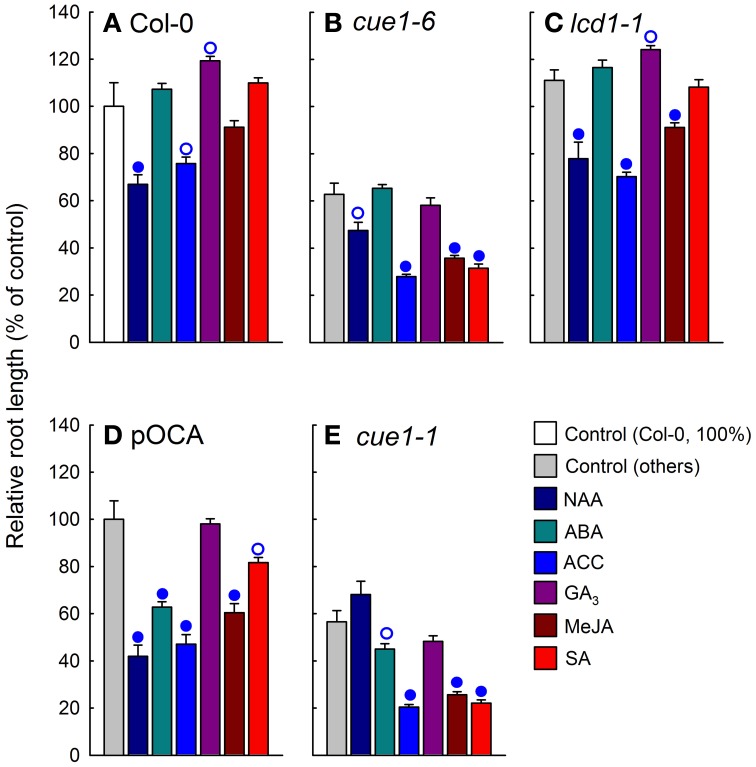
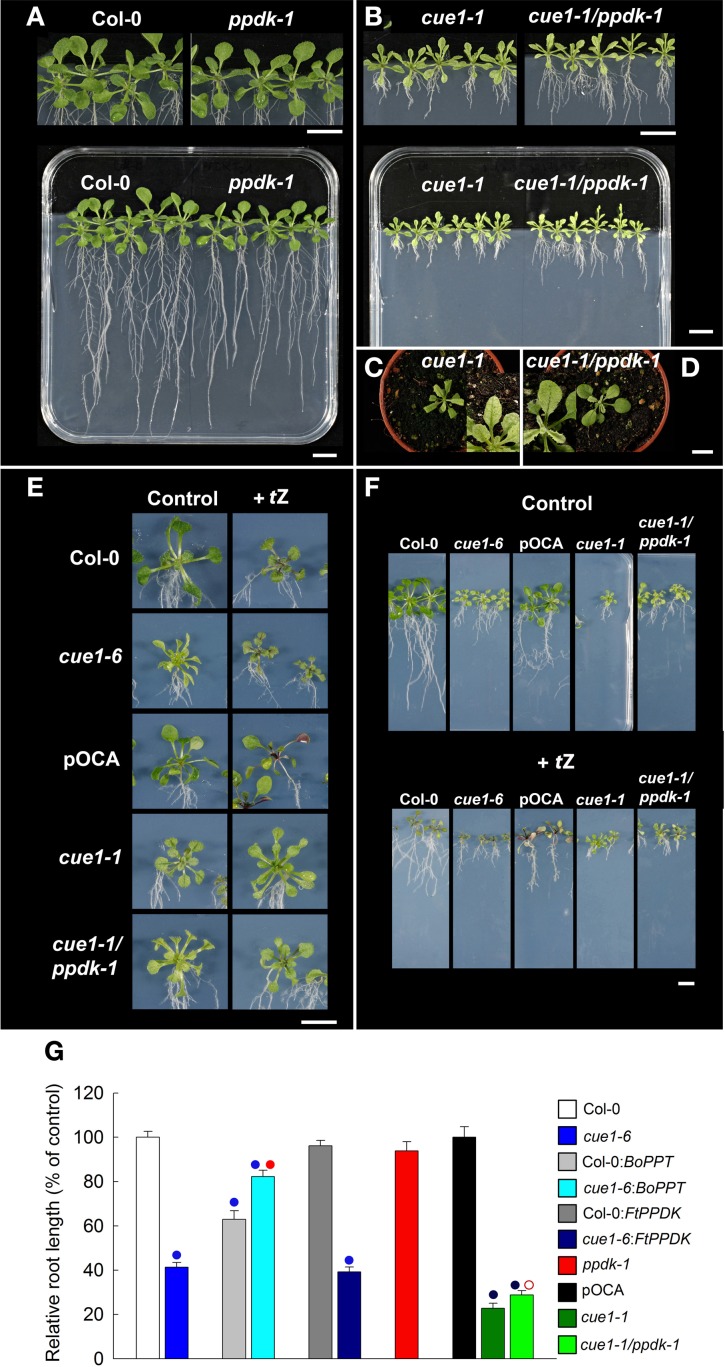
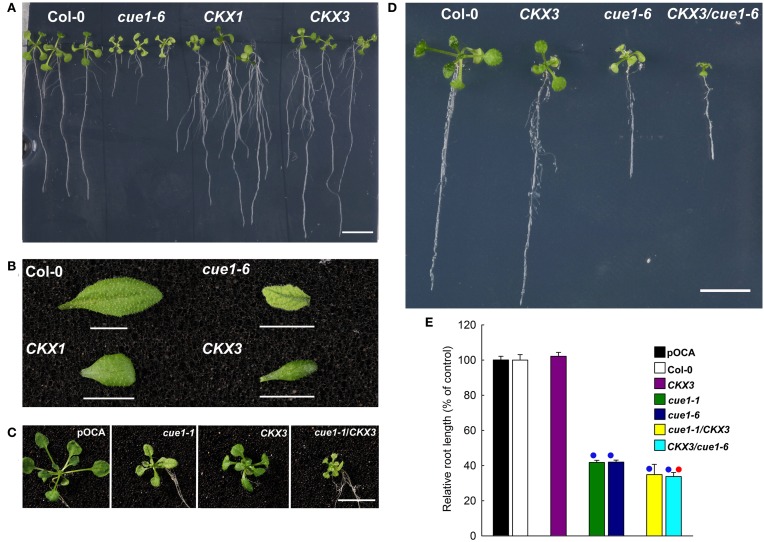
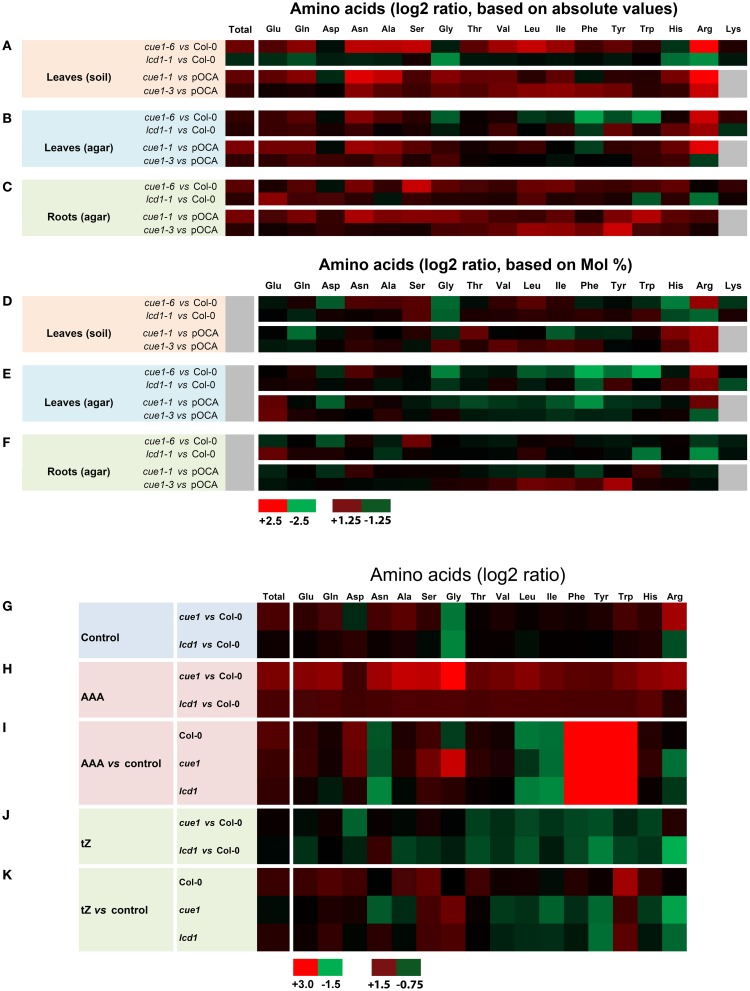
Phosphoenolpyruvate (PEP) serves not only as a high energy carbon compound in glycolysis, but it acts also as precursor for plastidial anabolic sequences like the shikimate pathway, which produces aromatic amino acids (AAA) and subsequently secondary plant products. After conversion to pyruvate, PEP can also enter de novo fatty acid biosynthesis, the synthesis of branched-chain amino acids, and the non-mevalonate way of isoprenoid production. As PEP cannot be generated by glycolysis in chloroplasts and a variety of non-green plastids, it has to be imported from the cytosol by a phosphate translocator (PT) specific for PEP (PPT). A loss of function of PPT1 in Arabidopsis thaliana results in the chlorophyll a/b binding protein underexpressed1 (cue1) mutant, which is characterized by reticulate leaves and stunted roots. Here we dissect the shoot- and root phenotypes, and also address the question whether or not long distance signaling by metabolites is involved in the perturbed mesophyll development of cue1. Reverse grafting experiments showed that the shoot- and root phenotypes develop independently from each other, ruling out long distance metabolite signaling. The leaf phenotype could be transiently modified even in mature leaves, e.g. by an inducible PPT1RNAi approach or by feeding AAA, the cytokinin trans-zeatin (tZ), or the putative signaling molecule dehydrodiconiferyl alcohol glucoside (DCG). Hormones, such as auxins, abscisic acid, gibberellic acid, ethylene, methyl jasmonate, and salicylic acid did not rescue the cue1 leaf phenotype. The low cell density1 (lcd1) mutant shares the reticulate leaf-, but not the stunted root phenotype with cue1. It could neither be rescued by AAA nor by tZ. In contrast, tZ and AAA further inhibited root growth both in cue1 and wild-type plants. Based on our results, we propose a model that PPT1 acts as a net importer of PEP into chloroplast, but as an overflow valve and hence exporter in root plastids.
磷酸烯醇丙酮酸(PEP)不仅是糖酵解过程中的高能碳化合物,而且还是质体合成途径的前体,如莽草酸途径,可产生芳香族氨基酸(AAA)和随后的次级植物产物。PEP 转化为丙酮酸后,也可以进入从头脂肪酸生物合成、支链氨基酸合成和非甲羟戊酸途径的异戊二烯生产。由于叶绿体和各种非绿色质体中不能通过糖酵解产生 PEP,因此必须通过特定于 PEP 的磷酸转运蛋白(PPT)从细胞质中输入。拟南芥中 PPT1 的功能丧失导致叶绿素 a/b 结合蛋白表达不足 1(cue1)突变体,其特征为网状叶片和根短。在这里,我们剖析了地上部和根部表型,还解决了代谢物是否参与 cue1 中受损的叶肉发育的长距离信号问题。反向嫁接实验表明,地上部和根部表型彼此独立发育,排除了长距离代谢物信号。即使在成熟叶片中,也可以暂时改变叶片表型,例如通过诱导性 PPT1RNAi 方法或通过饲喂 AAA、细胞分裂素玉米素(tZ)或假定的信号分子去氢二肉桂醇葡萄糖苷(DCG)。激素,如生长素、脱落酸、赤霉素、乙烯、茉莉酸甲酯和水杨酸,不能挽救 cue1 叶片表型。低细胞密度 1(lcd1)突变体与 cue1 共享网状叶片表型,但不具有短根表型。它既不能被 AAA 也不能被 tZ 挽救。相反,tZ 和 AAA 进一步抑制 cue1 和野生型植物的根生长。基于我们的结果,我们提出了一个模型,即 PPT1 作为 PEP 进入叶绿体的净输入器,但作为根质体中的溢流阀和出口器。