Castro-Diaz Nathaly, Ecco Gabriela, Coluccio Andrea, Kapopoulou Adamandia, Yazdanpanah Benyamin, Friedli Marc, Duc Julien, Jang Suk Min, Turelli Priscilla, Trono Didier
School of Life Sciences, Ecole Polytechnique Fédérale de Lausanne (EPFL), 1015 Lausanne, Switzerland.
Genes Dev. 2014 Jul 1;28(13):1397-409. doi: 10.1101/gad.241661.114. Epub 2014 Jun 17.
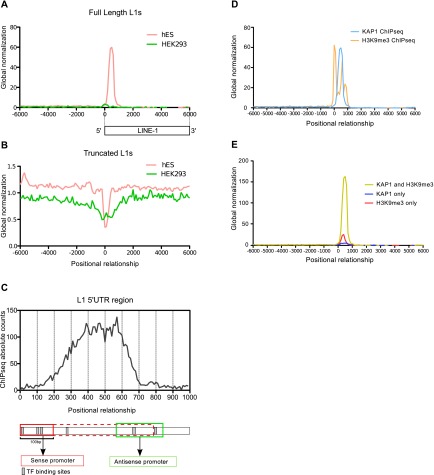
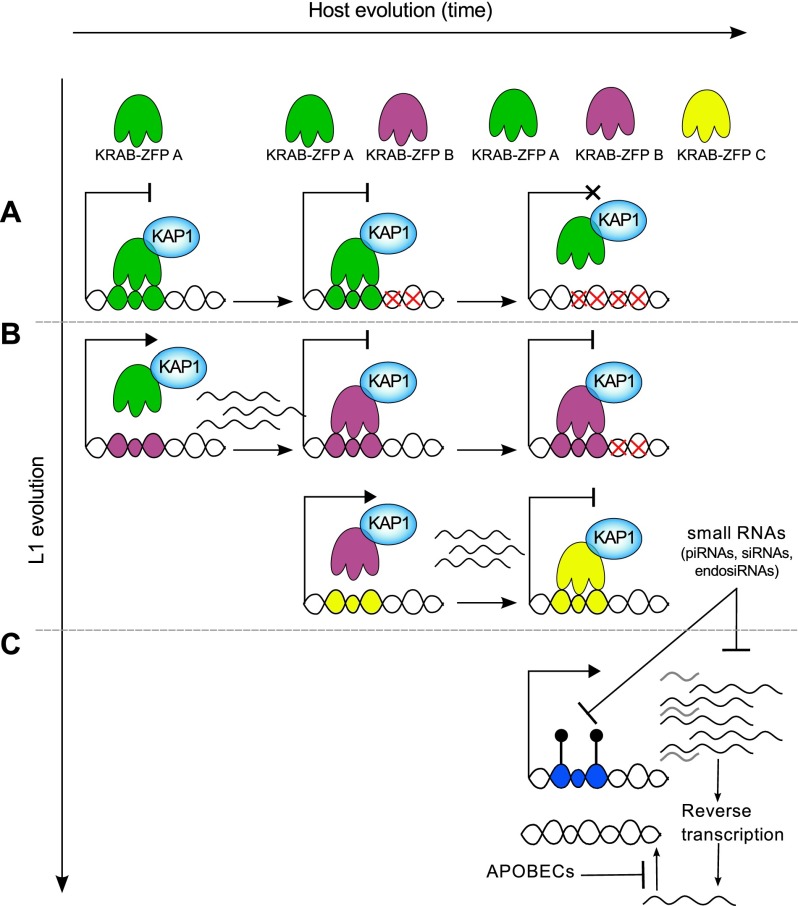
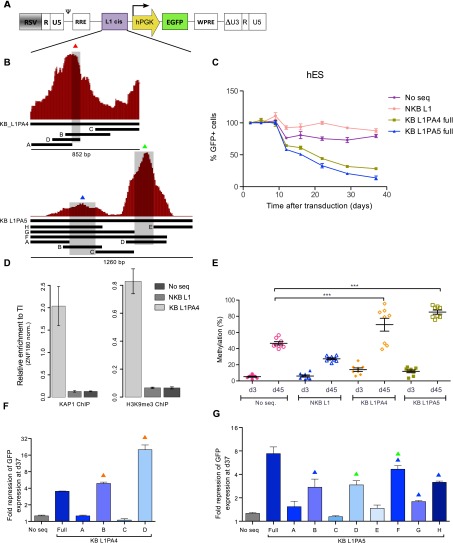
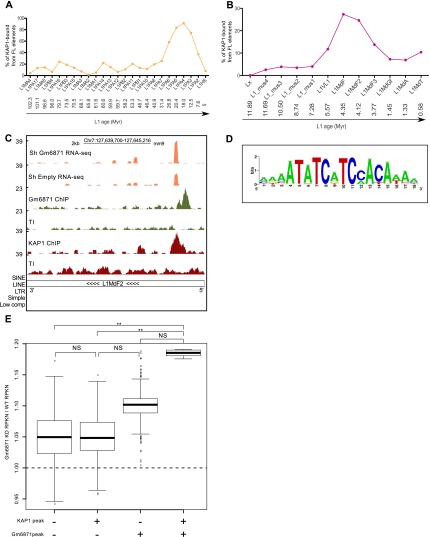
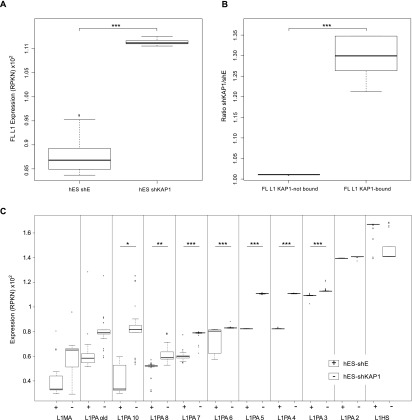
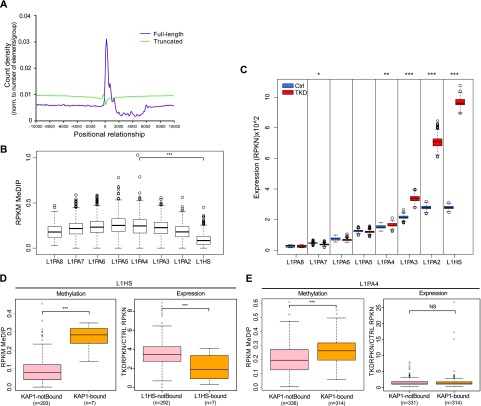
Mobile elements are important evolutionary forces that challenge genomic integrity. Long interspersed element-1 (L1, also known as LINE-1) is the only autonomous transposon still active in the human genome. It displays an unusual pattern of evolution, with, at any given time, a single active L1 lineage amplifying to thousands of copies before getting replaced by a new lineage, likely under pressure of host restriction factors, which act notably by silencing L1 expression during early embryogenesis. Here, we demonstrate that in human embryonic stem (hES) cells, KAP1 (KRAB [Krüppel-associated box domain]-associated protein 1), the master cofactor of KRAB-containing zinc finger proteins (KRAB-ZFPs) previously implicated in the restriction of endogenous retroviruses, represses a discrete subset of L1 lineages predicted to have entered the ancestral genome between 26.8 million and 7.6 million years ago. In mice, we documented a similar chronologically conditioned pattern, albeit with a much contracted time scale. We could further identify an L1-binding KRAB-ZFP, suggesting that this rapidly evolving protein family is more globally responsible for L1 recognition. KAP1 knockdown in hES cells induced the expression of KAP1-bound L1 elements, but their younger, human-specific counterparts (L1Hs) were unaffected. Instead, they were stimulated by depleting DNA methyltransferases, consistent with recent evidence demonstrating that the PIWI-piRNA (PIWI-interacting RNA) pathway regulates L1Hs in hES cells. Altogether, these data indicate that the early embryonic control of L1 is an evolutionarily dynamic process and support a model in which newly emerged lineages are first suppressed by DNA methylation-inducing small RNA-based mechanisms before KAP1-recruiting protein repressors are selected.
移动元件是挑战基因组完整性的重要进化力量。长散在核元件1(L1,也称为LINE-1)是唯一仍在人类基因组中活跃的自主转座子。它呈现出一种不同寻常的进化模式,在任何给定时间,单个活跃的L1谱系会扩增到数千个拷贝,然后被一个新的谱系所取代,这可能是在宿主限制因子的压力下发生的,宿主限制因子在早期胚胎发育过程中主要通过沉默L1表达来发挥作用。在这里,我们证明,在人类胚胎干细胞(hES)中,KAP1(含KRAB[克氏锥虫相关盒结构域]相关蛋白1)是先前与内源性逆转录病毒限制有关的含KRAB锌指蛋白(KRAB-ZFPs)的主要辅助因子,它抑制了一个离散的L1谱系子集,这些谱系预计在2680万至760万年前进入祖先基因组。在小鼠中,我们记录了一种类似的按时间顺序排列的模式,尽管时间尺度要小得多。我们还能进一步鉴定出一种与L1结合的KRAB-ZFP,这表明这个快速进化的蛋白家族在更广泛的层面上负责L1的识别。在hES细胞中敲低KAP1会诱导与KAP1结合的L1元件的表达,但它们较年轻的、人类特有的对应物(L1Hs)不受影响。相反,它们会因DNA甲基转移酶的缺失而受到刺激,这与最近的证据一致,该证据表明PIWI- piRNA(PIWI相互作用RNA)途径在hES细胞中调节L1Hs。总之,这些数据表明L1的早期胚胎控制是一个进化上动态的过程,并支持一种模型,即新出现的谱系首先被基于DNA甲基化诱导的小RNA机制所抑制,然后才选择招募KAP1的蛋白抑制因子。