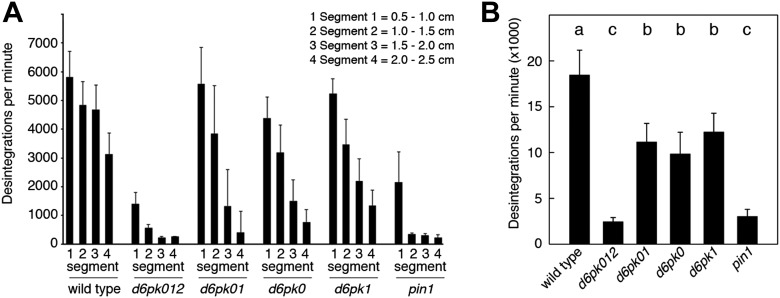
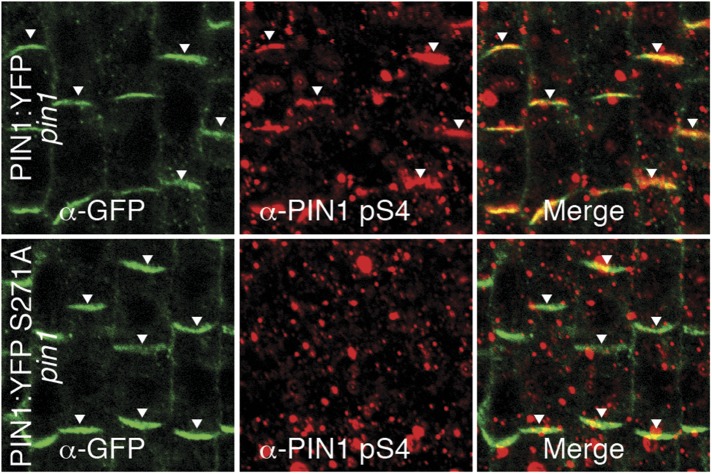
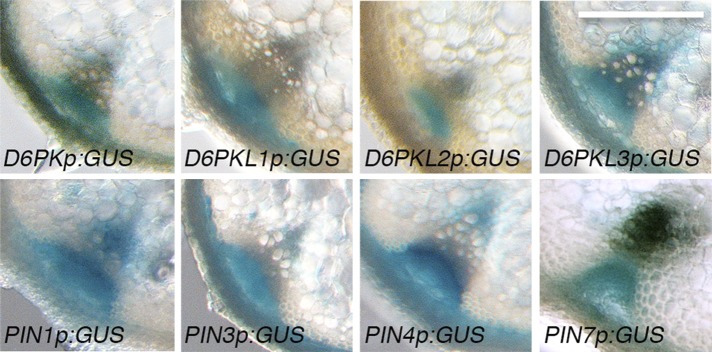
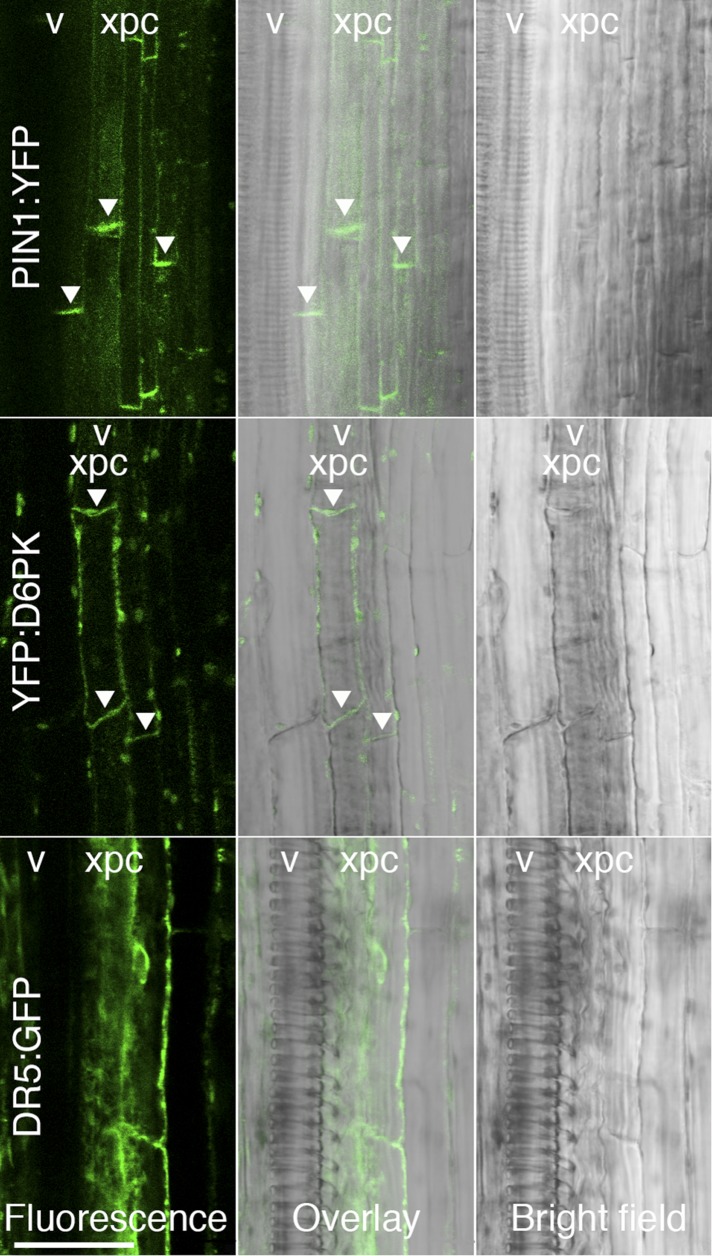
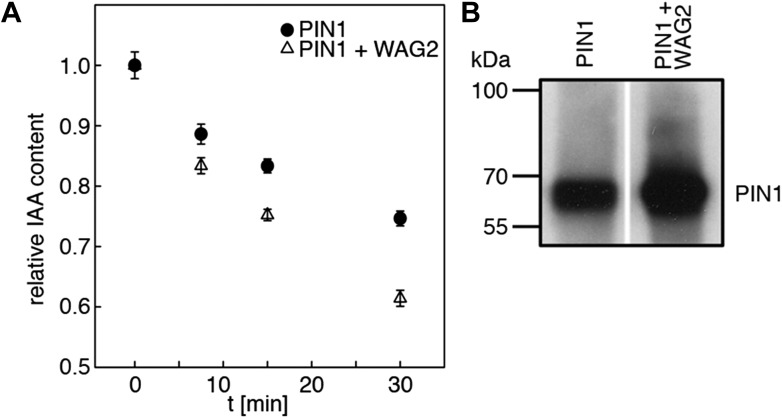
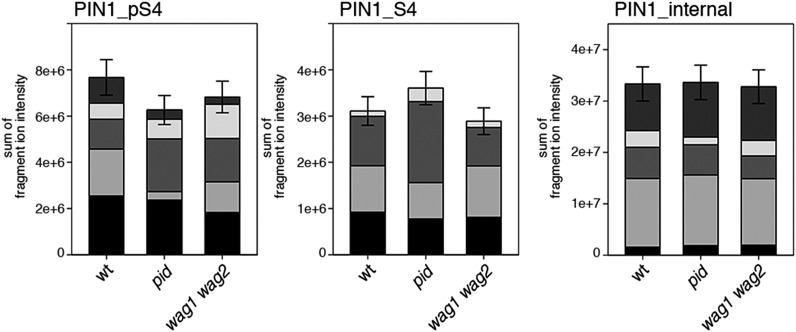
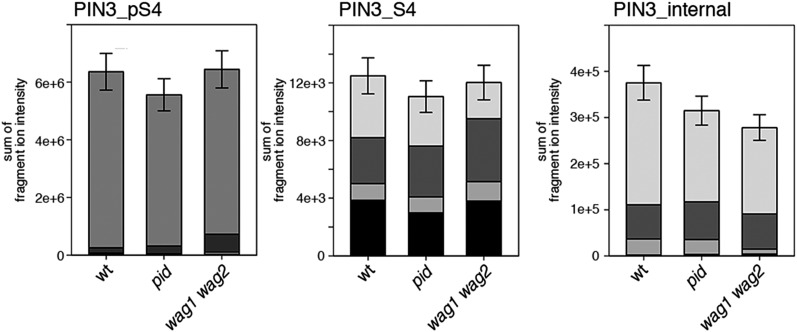
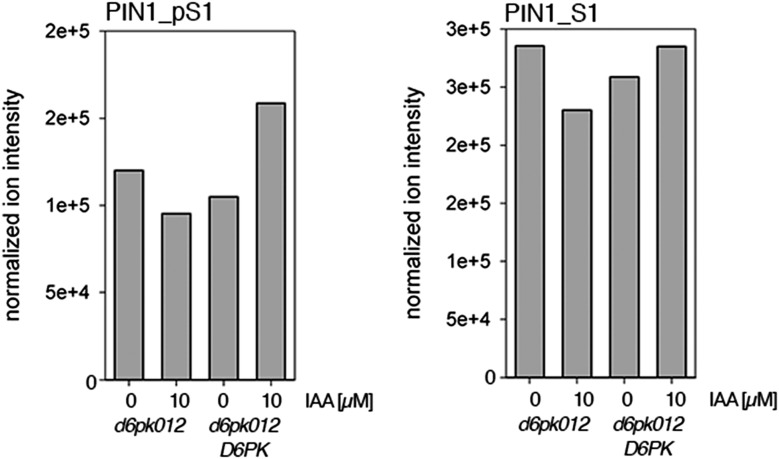
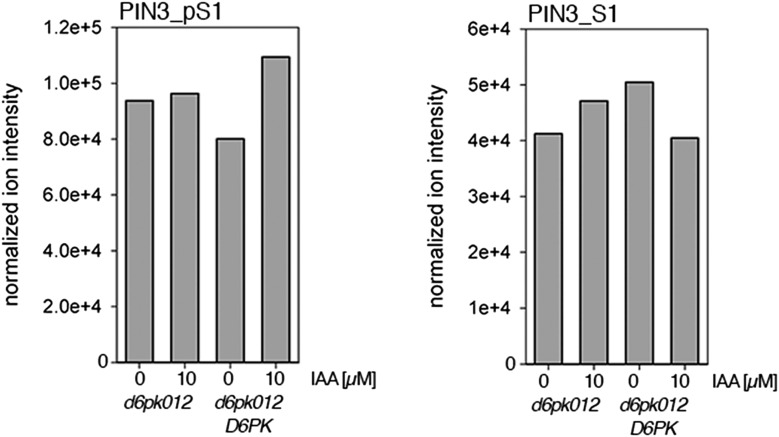
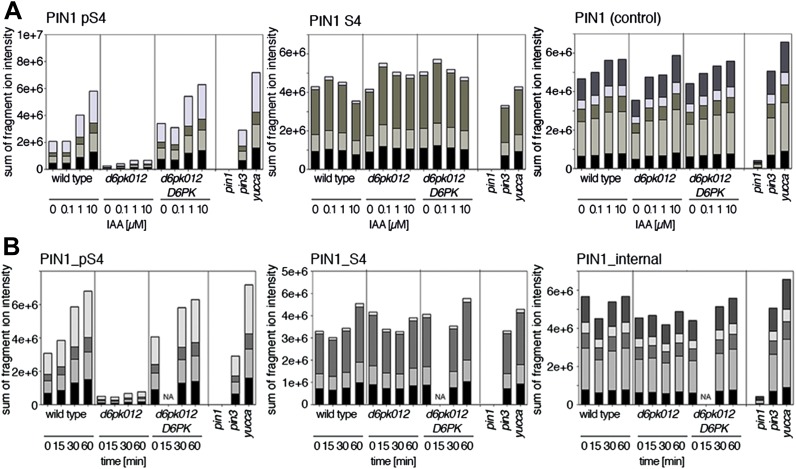
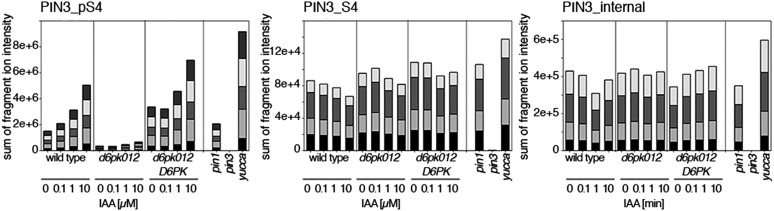
Zourelidou Melina, Absmanner Birgit, Weller Benjamin, Barbosa Inês C R, Willige Björn C, Fastner Astrid, Streit Verena, Port Sarah A, Colcombet Jean, de la Fuente van Bentem Sergio, Hirt Heribert, Kuster Bernhard, Schulze Waltraud X, Hammes Ulrich Z, Schwechheimer Claus
Department of Plant Systems Biology, Technische Universität München, Freising, Germany.
Department of Cell Biology and Plant Biochemistry, Universität Regensburg, Regensburg, Germany.
Elife. 2014 Jun 19;3:e02860. doi: 10.7554/eLife.02860.
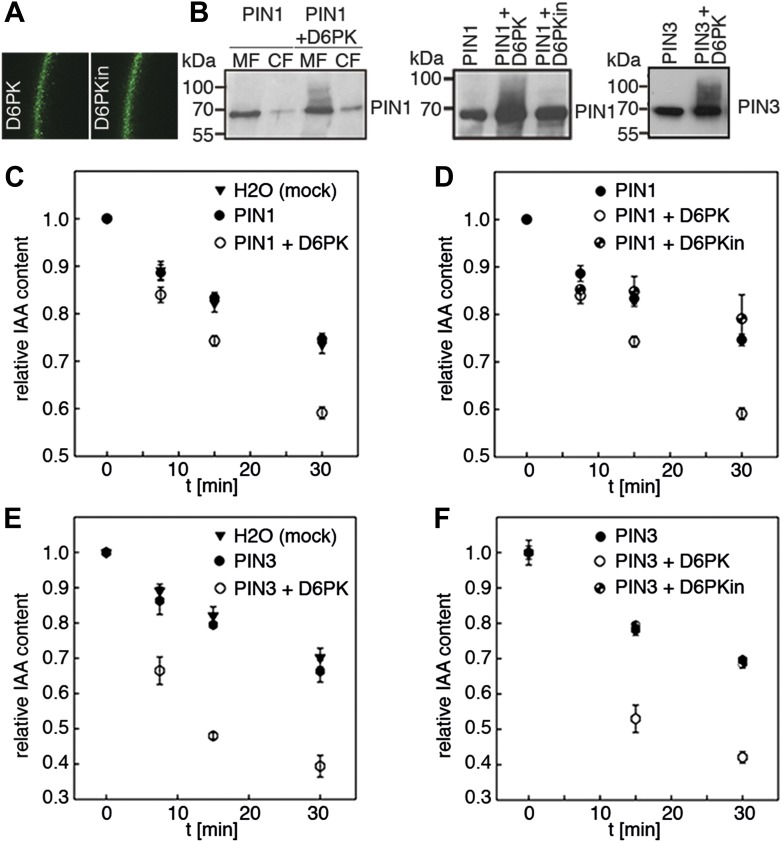
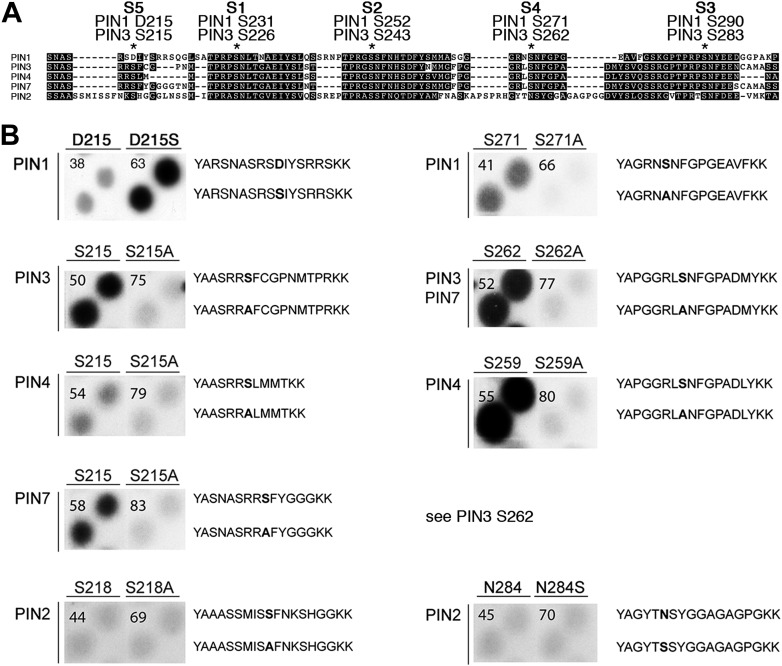
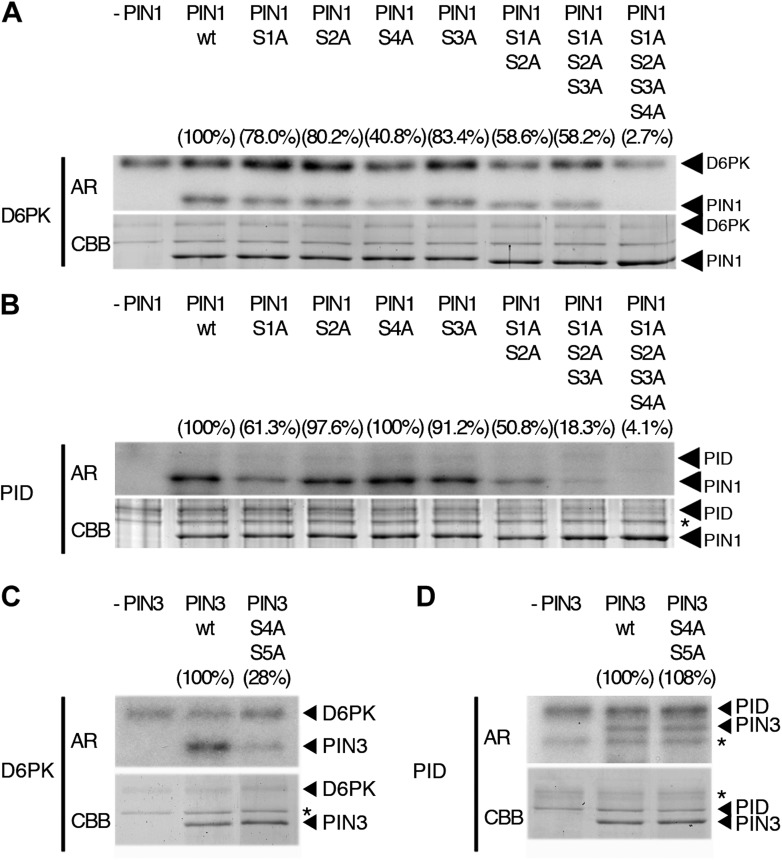
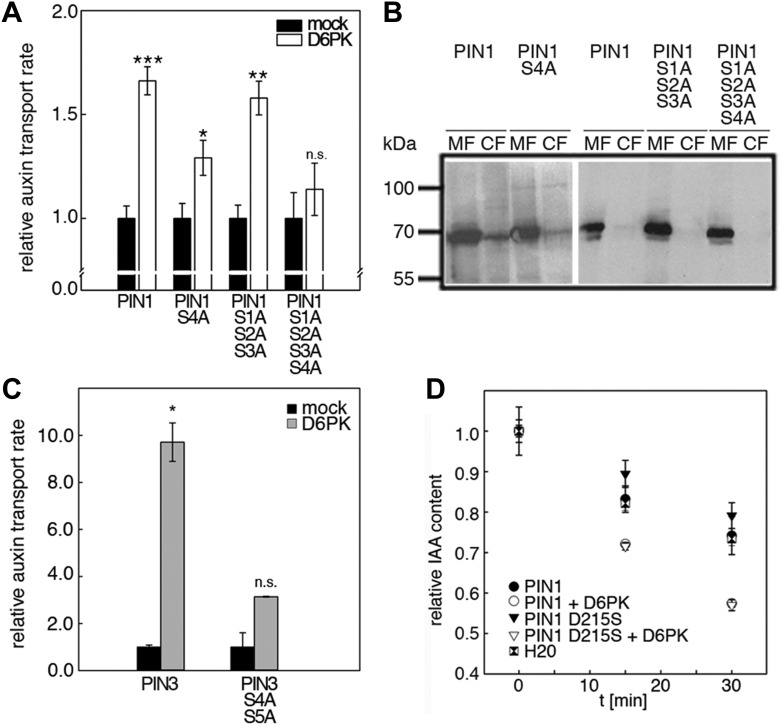
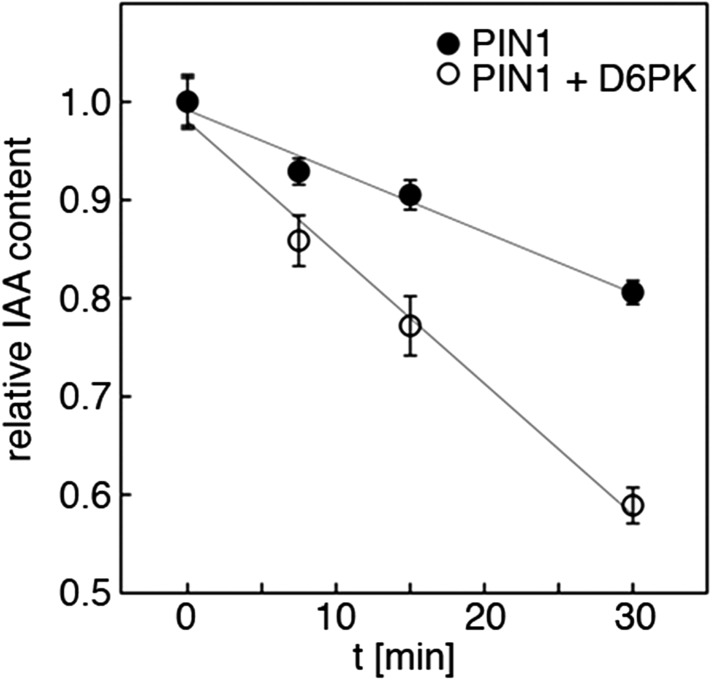
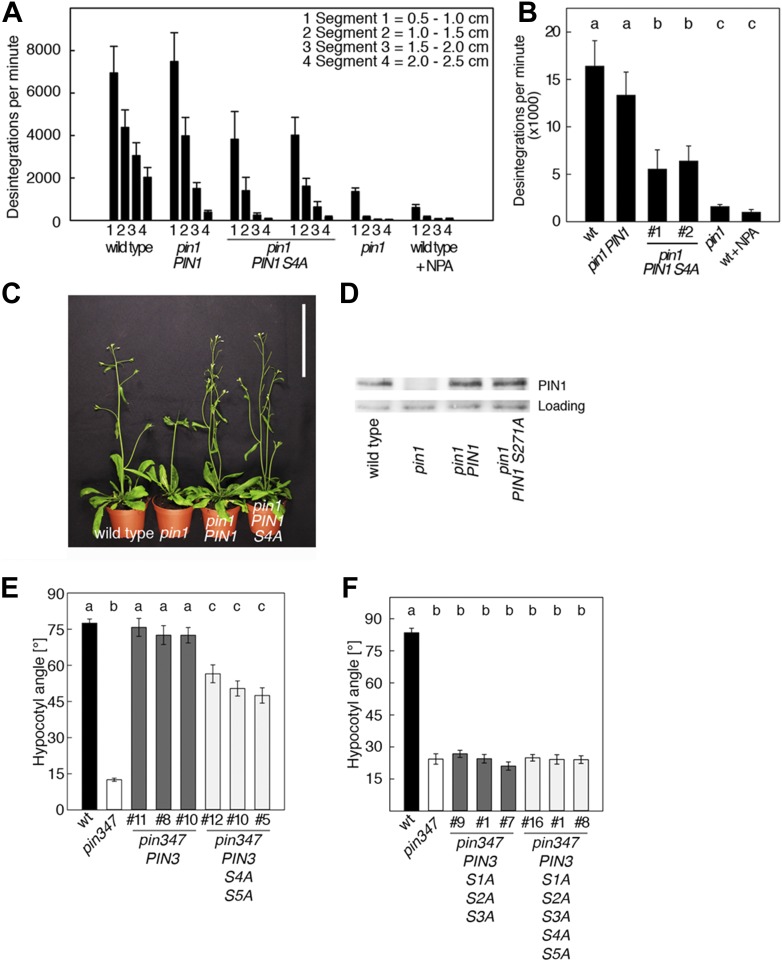
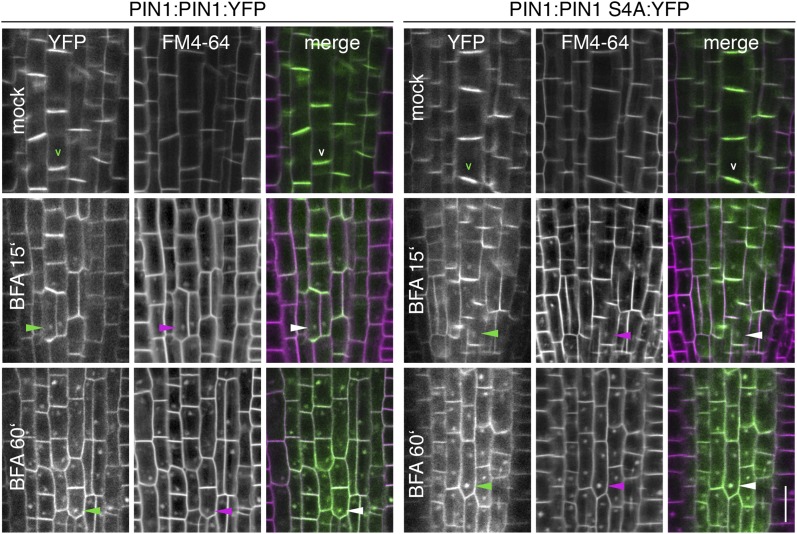
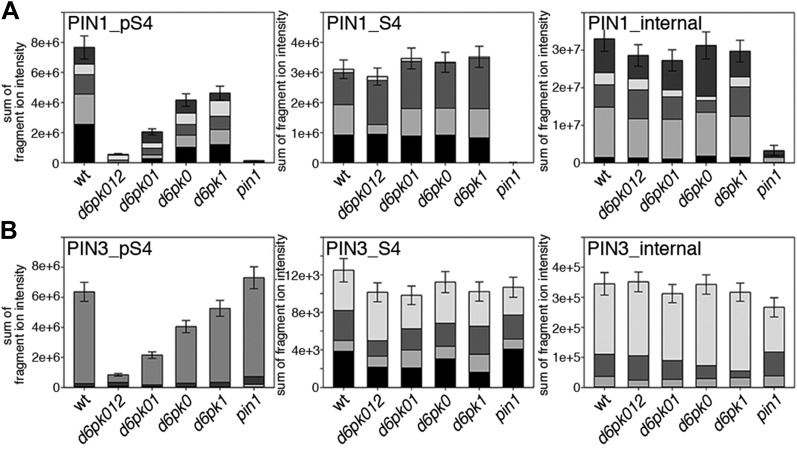
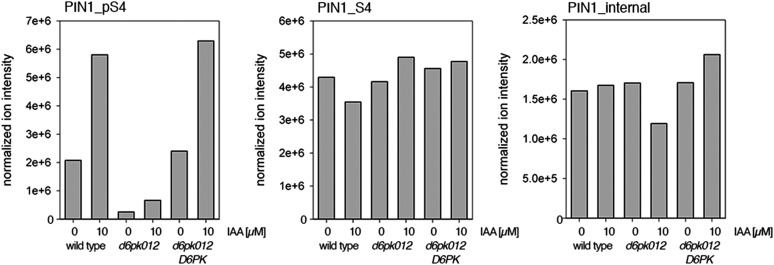
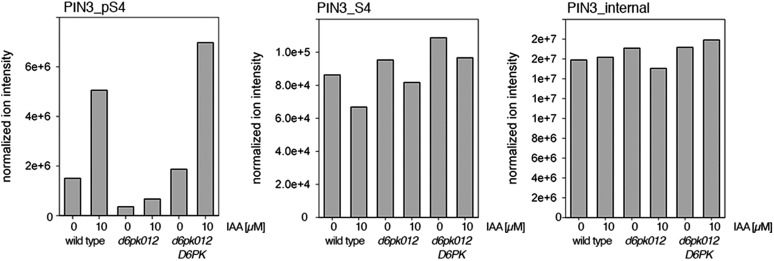
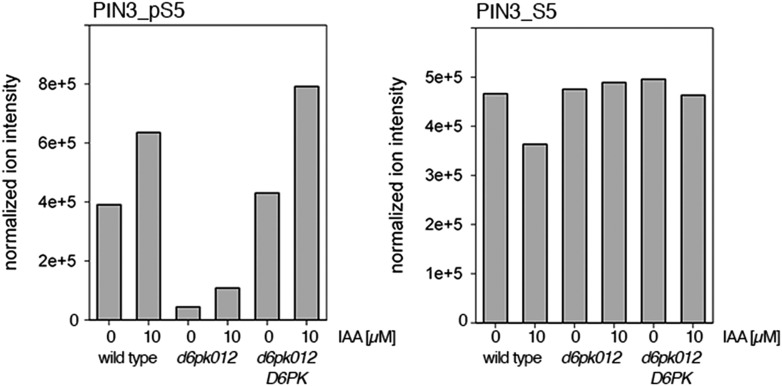
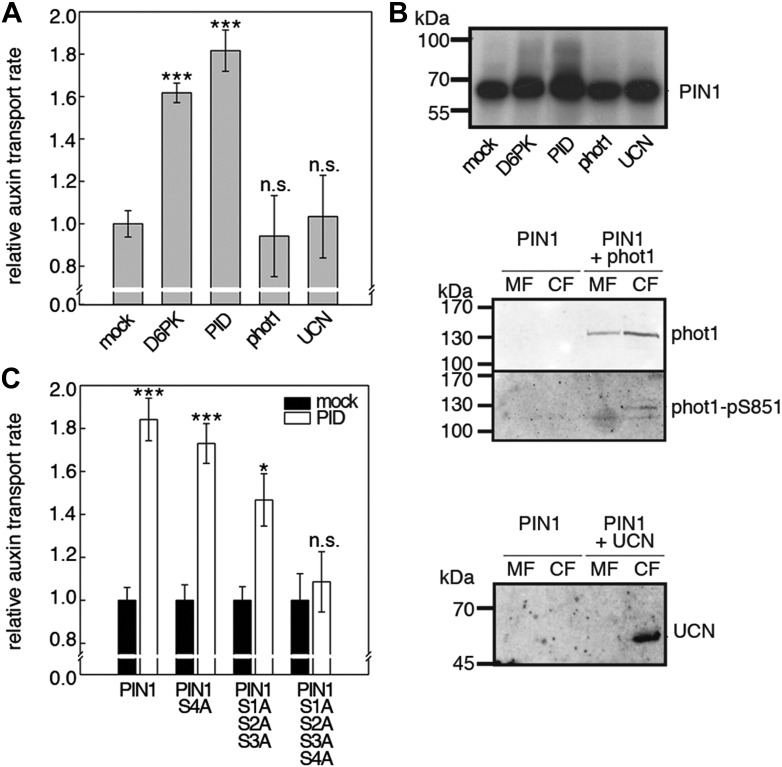
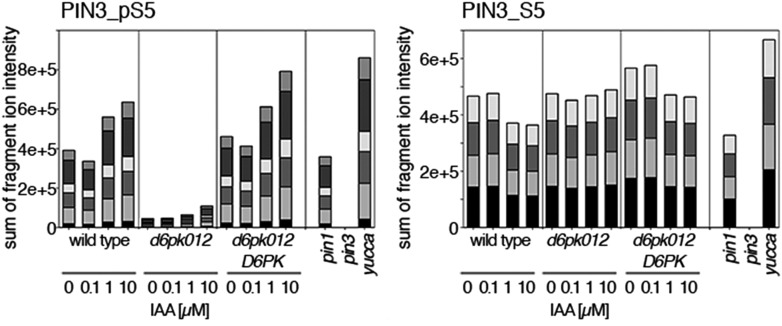
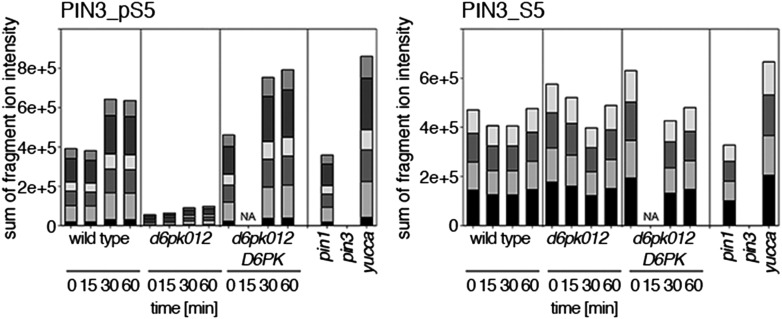
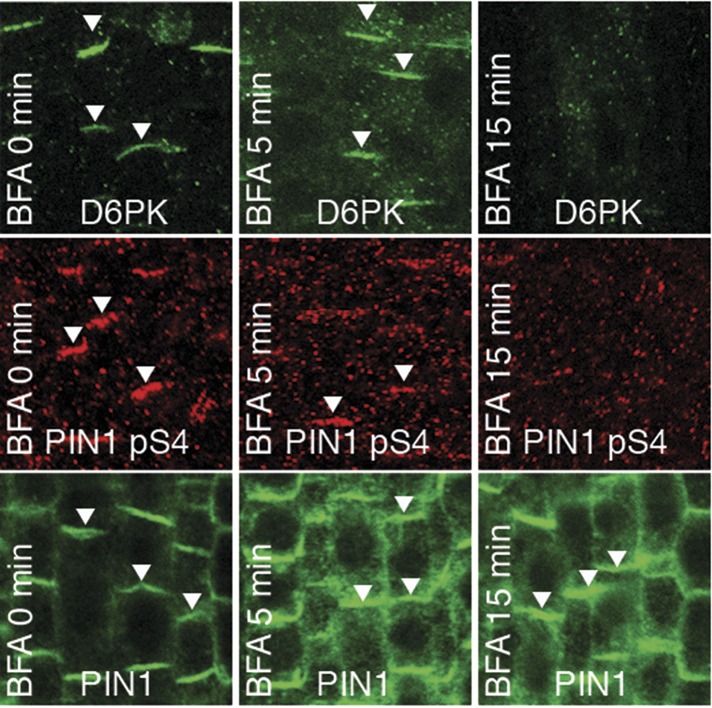
The development and morphology of vascular plants is critically determined by synthesis and proper distribution of the phytohormone auxin. The directed cell-to-cell distribution of auxin is achieved through a system of auxin influx and efflux transporters. PIN-FORMED (PIN) proteins are proposed auxin efflux transporters, and auxin fluxes can seemingly be predicted based on the--in many cells--asymmetric plasma membrane distribution of PINs. Here, we show in a heterologous Xenopus oocyte system as well as in Arabidopsis thaliana inflorescence stems that PIN-mediated auxin transport is directly activated by D6 PROTEIN KINASE (D6PK) and PINOID (PID)/WAG kinases of the Arabidopsis AGCVIII kinase family. At the same time, we reveal that D6PKs and PID have differential phosphosite preferences. Our study suggests that PIN activation by protein kinases is a crucial component of auxin transport control that must be taken into account to understand auxin distribution within the plant.
维管植物的发育和形态在很大程度上取决于植物激素生长素的合成和适当分布。生长素在细胞间的定向分布是通过生长素流入和流出转运体系统实现的。PIN形成蛋白(PIN)被认为是生长素流出转运体,并且在许多细胞中,生长素流似乎可以基于PIN在质膜上的不对称分布来预测。在这里,我们在异源非洲爪蟾卵母细胞系统以及拟南芥花序茎中表明,PIN介导的生长素转运直接由拟南芥AGCVIII激酶家族的D6蛋白激酶(D6PK)和类PID激酶(PID)/WAG激酶激活。同时,我们发现D6PK和PID具有不同的磷酸化位点偏好。我们的研究表明,蛋白激酶对PIN的激活是生长素运输控制的关键组成部分,要了解植物体内生长素的分布,必须考虑这一点。