Morikawa Masayuki, Kino Katsuhito, Oyoshi Takanori, Suzuki Masayo, Kobayashi Takanobu, Miyazawa Hiroshi
Kagawa School of Pharmaceutical Sciences, Tokushima Bunri University, 1314-1 Shido, Sanuki, Kagawa 769-2193, Japan.
Faculty of Science, Department of Chemistry, Shizuoka University, 836 Ohya, Suruga, Shizuoka 422-8529, Japan.
Biomolecules. 2014 Feb 10;4(1):140-59. doi: 10.3390/biom4010140.
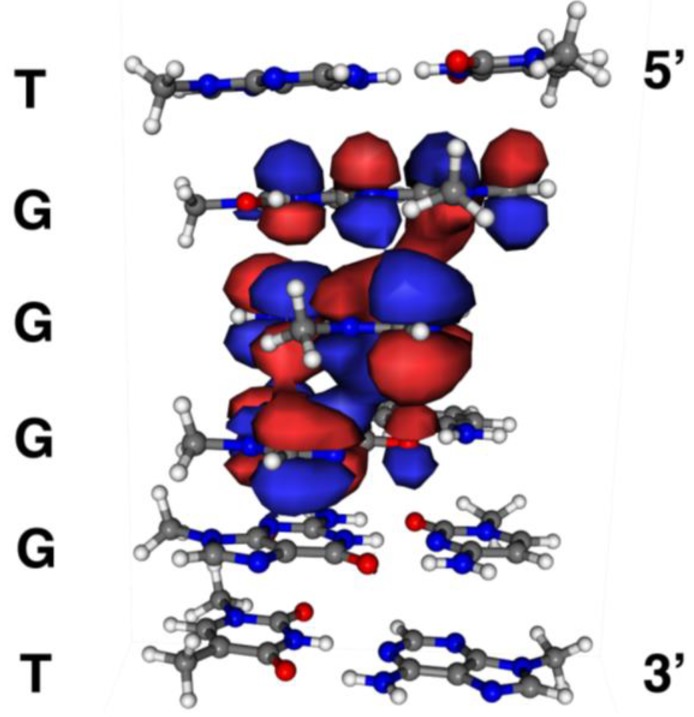
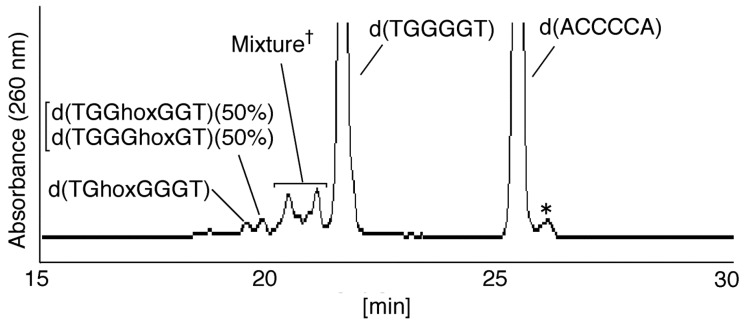
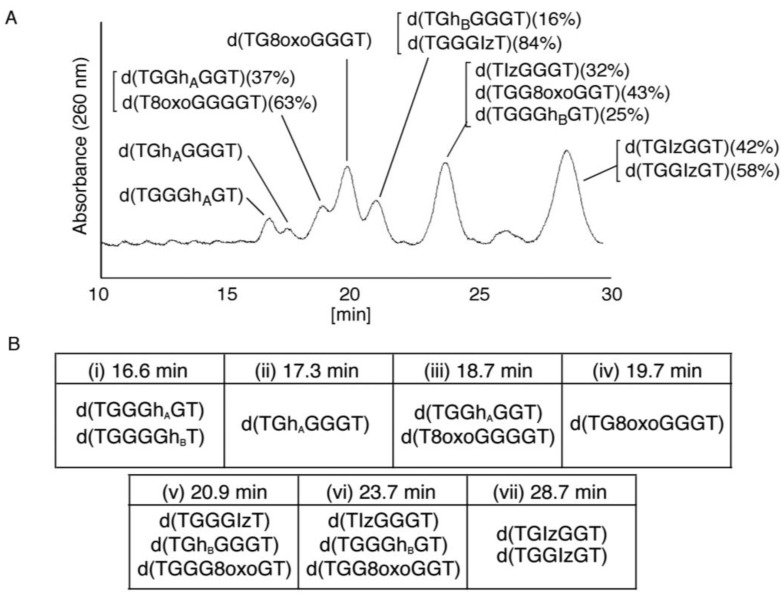
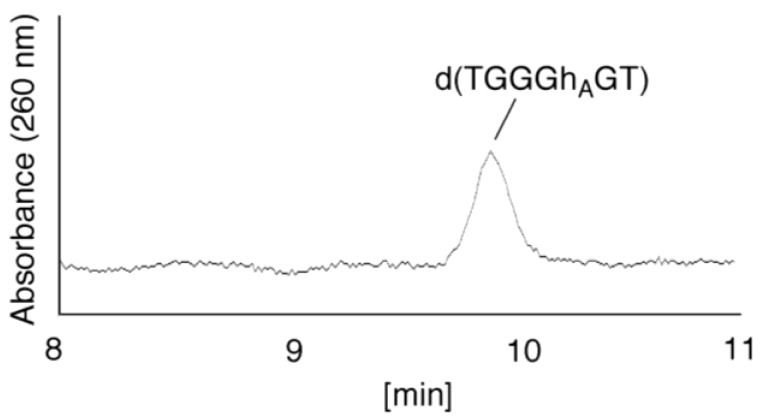
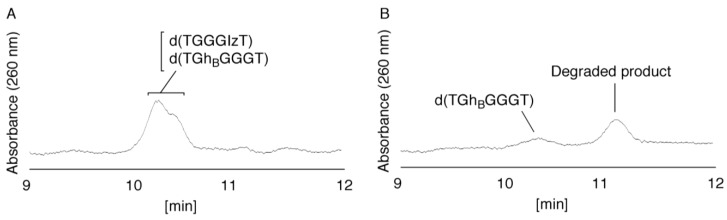
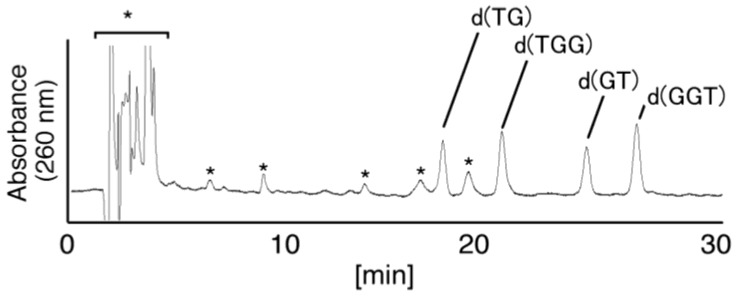
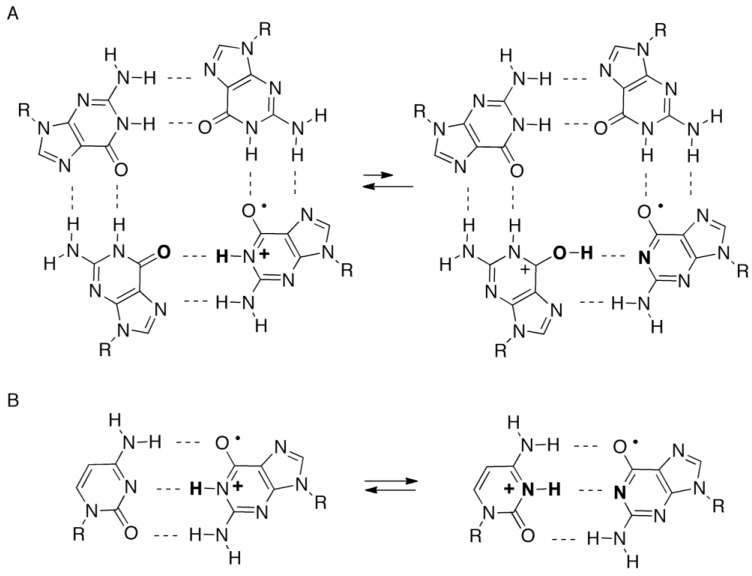
Guanine is the most easily oxidized among the four DNA bases, and some guanine-rich sequences can form quadruplex structures. In a previous study using 6-mer DNA d(TGGGGT), which is the shortest oligomer capable of forming quadruplex structures, we demonstrated that guanine oxidation products of quadruplex DNA differ from those of single-stranded DNA. Therefore, the hotooxidation products of double-stranded DNA (dsDNA) may also differ from that of quadruplex or single-stranded DNA, with the difference likely explaining the influence of DNA structures on guanine oxidation pathways. In this study, the guanine oxidation products of the dsDNA d(TGGGGT)/d(ACCCCA) were analyzed using HPLC and electrospray ionization-mass spectrometry (ESI-MS). As a result, the oxidation products in this dsDNA were identified as 2,5-diamino-4H-imidazol-4-one (Iz), 8-oxo-7,8-dihydroguanine (8oxoG), dehydroguanidinohydantoin (Ghox), and guanidinohydantoin (Gh). The major oxidation products in dsDNA were consistent with a combination of each major oxidation product observed in single-stranded and quadruplex DNA. We previously reported that the kinds of the oxidation products in single-stranded or quadruplex DNA depend on the ease of deprotonation of the guanine radical cation (G•+) at the N1 proton. Similarly, this mechanism was also involved in dsDNA. Deprotonation in dsDNA is easier than in quadruplex DNA and more difficult in single-stranded DNA, which can explain the formation of the four oxidation products in dsDNA.
鸟嘌呤是四种DNA碱基中最易被氧化的,一些富含鸟嘌呤的序列可形成四链体结构。在先前一项使用6聚体DNA d(TGGGGT)(能够形成四链体结构的最短寡聚物)的研究中,我们证明了四链体DNA的鸟嘌呤氧化产物与单链DNA的不同。因此,双链DNA(dsDNA)的光氧化产物可能也与四链体或单链DNA的不同,这种差异可能解释了DNA结构对鸟嘌呤氧化途径的影响。在本研究中,使用高效液相色谱法和电喷雾电离质谱法(ESI-MS)分析了dsDNA d(TGGGGT)/d(ACCCCA)的鸟嘌呤氧化产物。结果,该dsDNA中的氧化产物被鉴定为2,5-二氨基-4H-咪唑-4-酮(Iz)、8-氧代-7,8-二氢鸟嘌呤(8oxoG)、脱氢胍基乙内酰脲(Ghox)和胍基乙内酰脲(Gh)。dsDNA中的主要氧化产物与在单链和四链体DNA中观察到的每种主要氧化产物的组合一致。我们先前报道过,单链或四链体DNA中氧化产物的种类取决于鸟嘌呤自由基阳离子(G•+)在N1质子处去质子化的难易程度。同样,这种机制也在dsDNA中起作用。dsDNA中的去质子化比四链体DNA中更容易,比单链DNA中更难,这可以解释dsDNA中四种氧化产物的形成。