Lee Chiara, Yashiro Shoko, Dotson David L, Uzdavinys Povilas, Iwata So, Sansom Mark S P, von Ballmoos Christoph, Beckstein Oliver, Drew David, Cameron Alexander D
Division of Molecular Biosciences, Imperial College London, London SW7 2AZ, England, UK.
Division of Molecular Biosciences, Imperial College London, London SW7 2AZ, England, UK Membrane Protein Laboratory, Diamond Light Source, Harwell Science and Innovation Campus, Oxfordshire OX11 0DE, England, UK.
J Gen Physiol. 2014 Dec;144(6):529-44. doi: 10.1085/jgp.201411219.
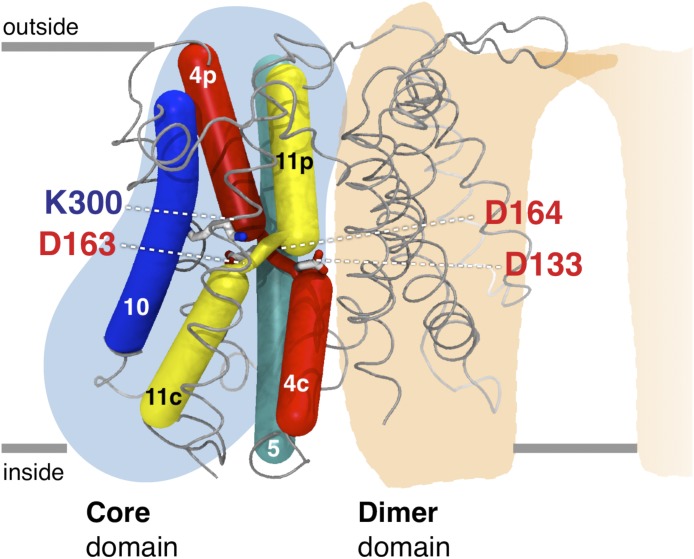
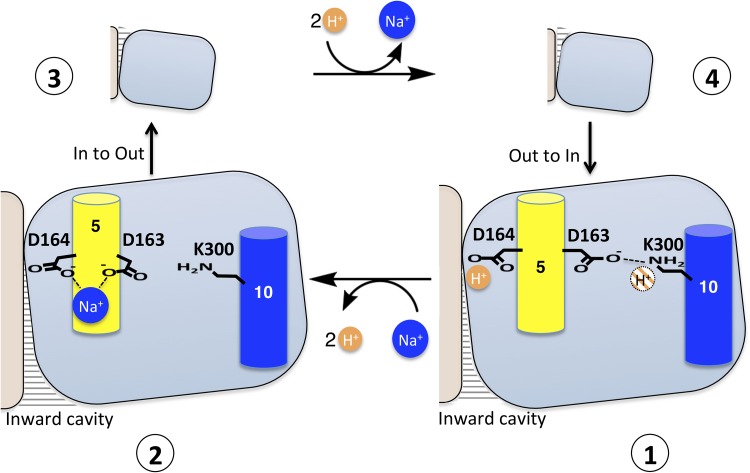
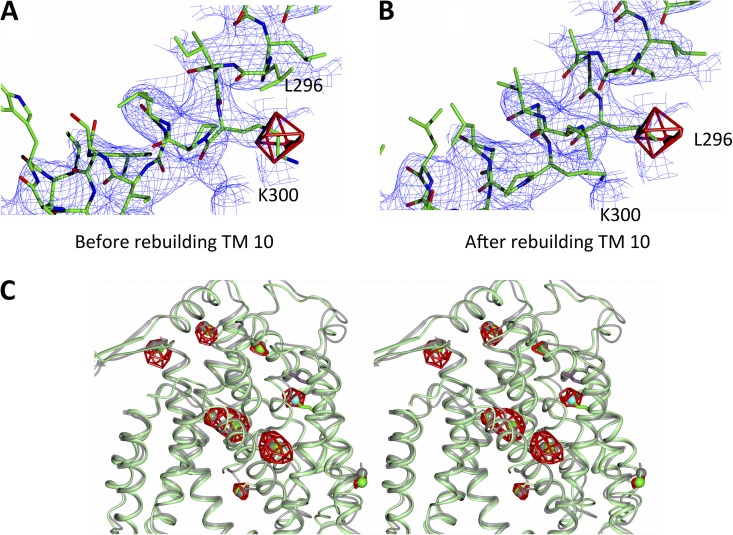
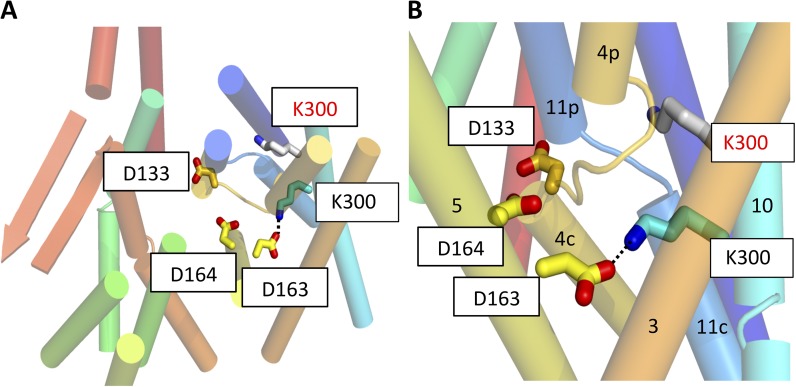
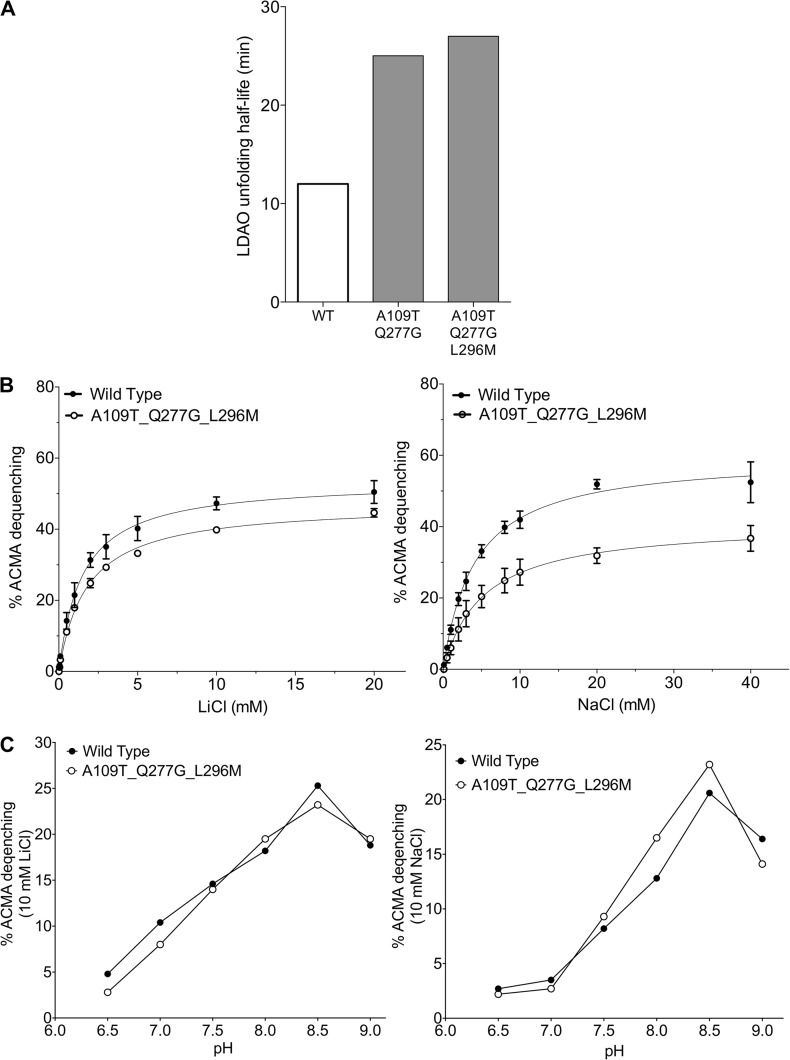
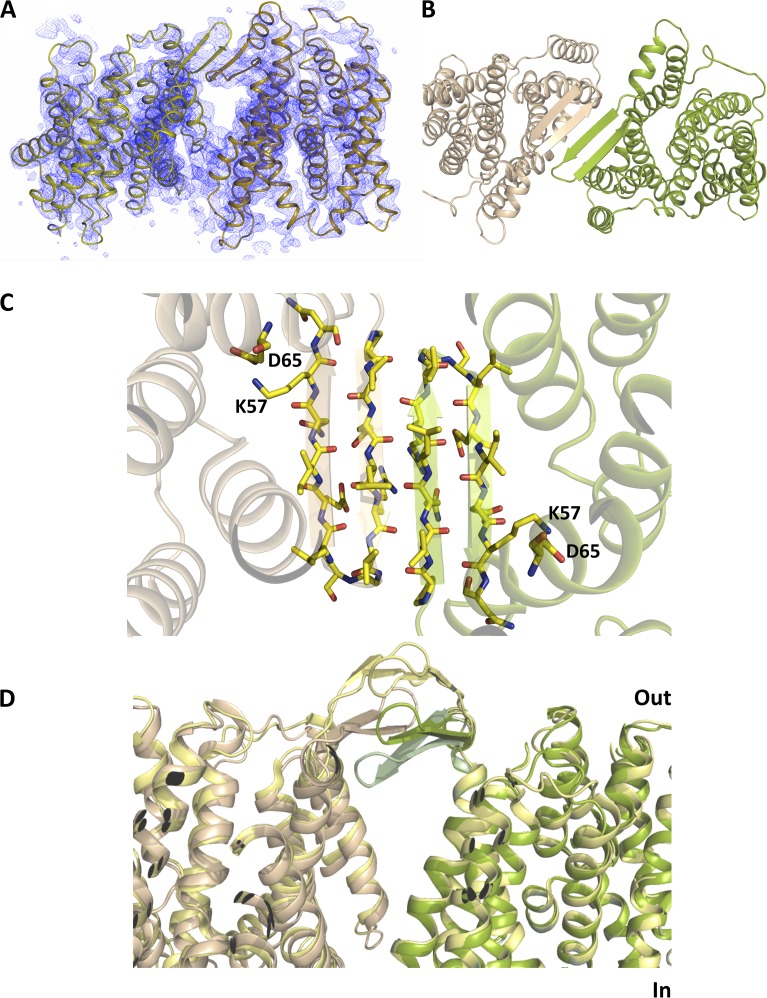
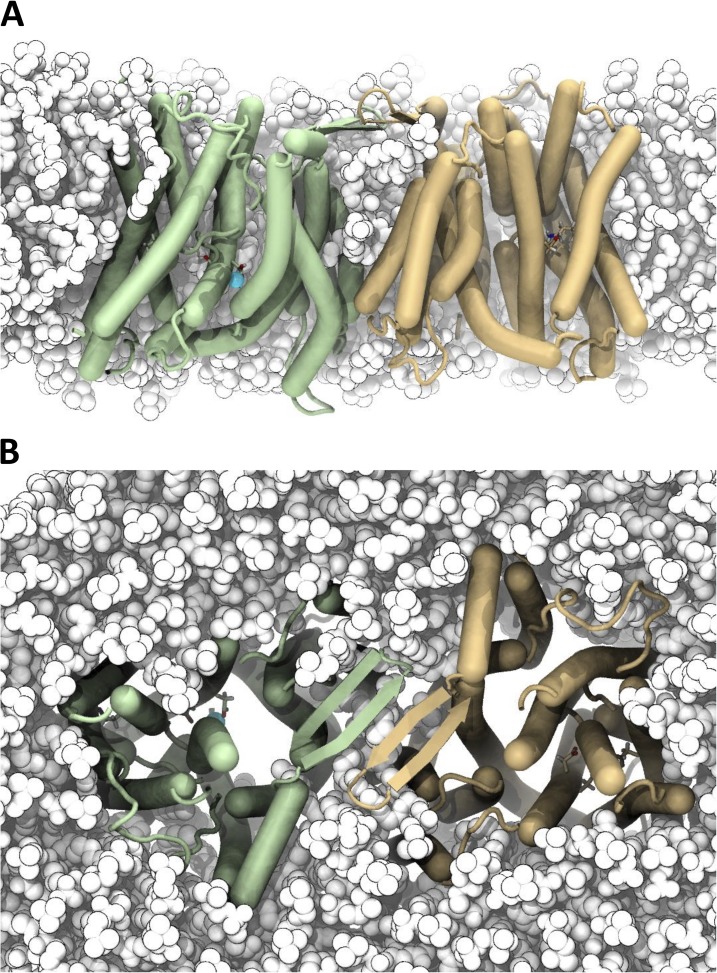
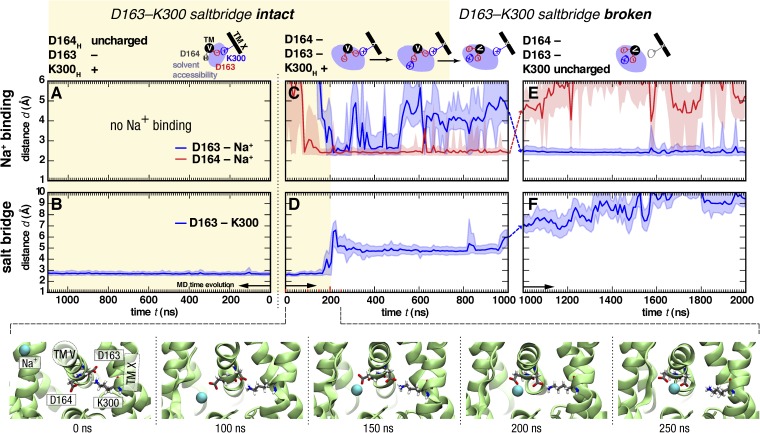
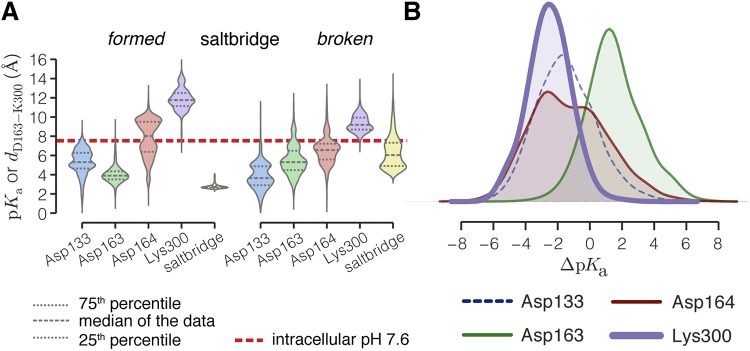
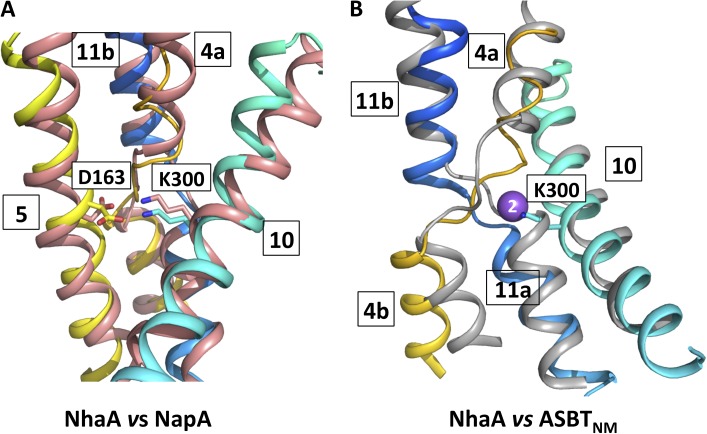
Sodium-proton antiporters rapidly exchange protons and sodium ions across the membrane to regulate intracellular pH, cell volume, and sodium concentration. How ion binding and release is coupled to the conformational changes associated with transport is not clear. Here, we report a crystal form of the prototypical sodium-proton antiporter NhaA from Escherichia coli in which the protein is seen as a dimer. In this new structure, we observe a salt bridge between an essential aspartic acid (Asp163) and a conserved lysine (Lys300). An equivalent salt bridge is present in the homologous transporter NapA, but not in the only other known crystal structure of NhaA, which provides the foundation of most existing structural models of electrogenic sodium-proton antiport. Molecular dynamics simulations show that the stability of the salt bridge is weakened by sodium ions binding to Asp164 and the neighboring Asp163. This suggests that the transport mechanism involves Asp163 switching between forming a salt bridge with Lys300 and interacting with the sodium ion. pKa calculations suggest that Asp163 is highly unlikely to be protonated when involved in the salt bridge. As it has been previously suggested that Asp163 is one of the two residues through which proton transport occurs, these results have clear implications to the current mechanistic models of sodium-proton antiport in NhaA.
钠-质子反向转运体可快速跨膜交换质子和钠离子,以调节细胞内pH值、细胞体积和钠浓度。目前尚不清楚离子的结合与释放是如何与转运相关的构象变化相耦合的。在此,我们报道了来自大肠杆菌的典型钠-质子反向转运体NhaA的一种晶体形态,其中该蛋白呈现为二聚体形式。在这个新结构中,我们观察到一个必需的天冬氨酸(Asp163)与一个保守的赖氨酸(Lys300)之间形成了一个盐桥。同源转运体NapA中也存在一个等效的盐桥,但在NhaA唯一已知的另一种晶体结构中不存在,而后者是大多数现有电生钠-质子反向转运结构模型的基础。分子动力学模拟表明,钠离子与Asp164及相邻的Asp163结合会削弱盐桥的稳定性。这表明转运机制涉及Asp163在与Lys300形成盐桥和与钠离子相互作用之间进行切换。pKa计算表明,当Asp163参与形成盐桥时,其被质子化的可能性极小。由于之前有人提出Asp163是质子转运所通过的两个残基之一,这些结果对目前NhaA中钠-质子反向转运的机制模型具有明确的启示。