Division of Molecular Biosciences, Imperial College London, London SW7 2AZ, UK.
Nature. 2013 Sep 26;501(7468):573-7. doi: 10.1038/nature12484. Epub 2013 Sep 1.
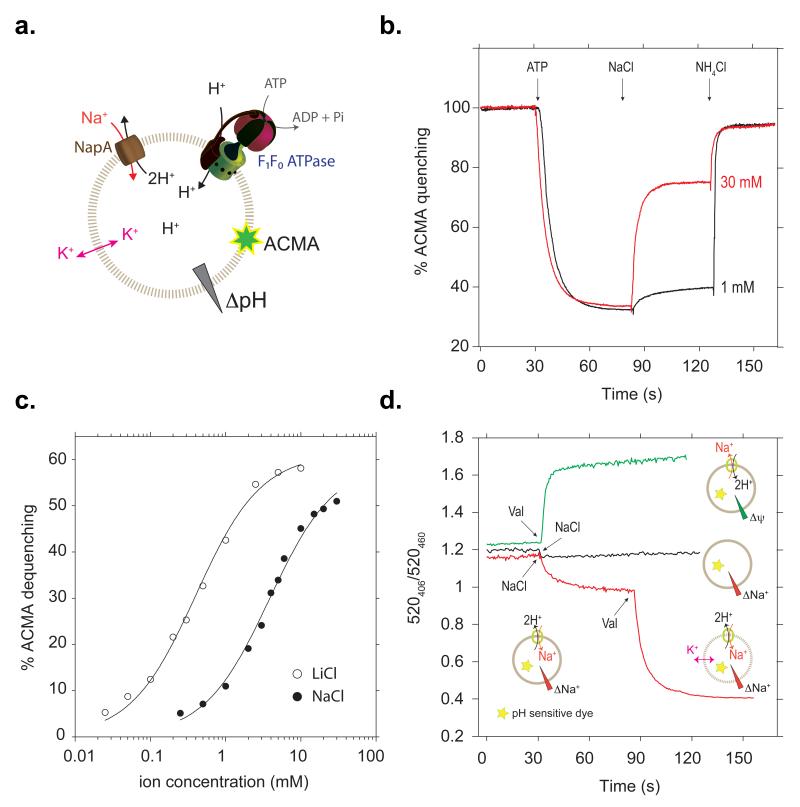
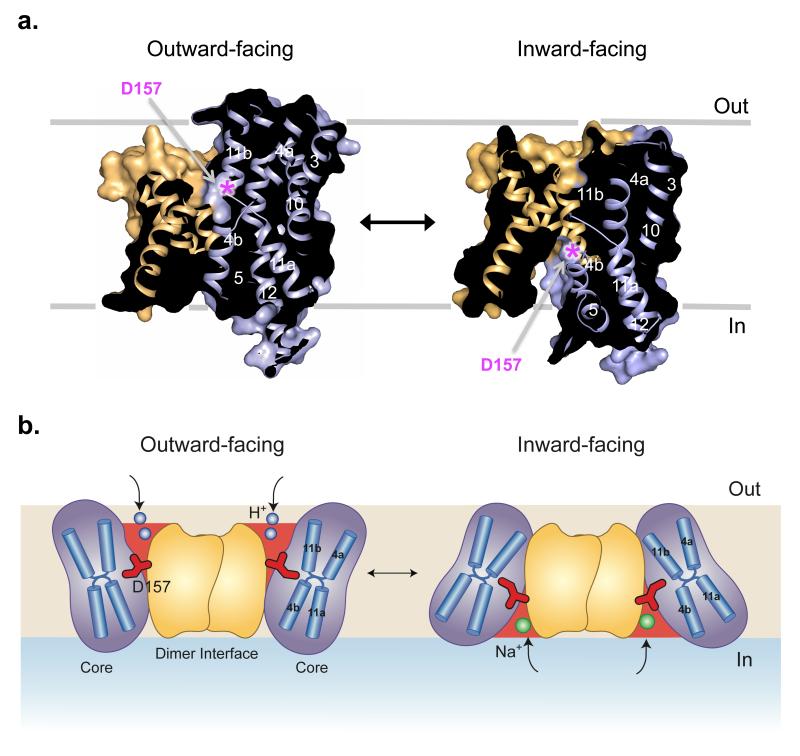
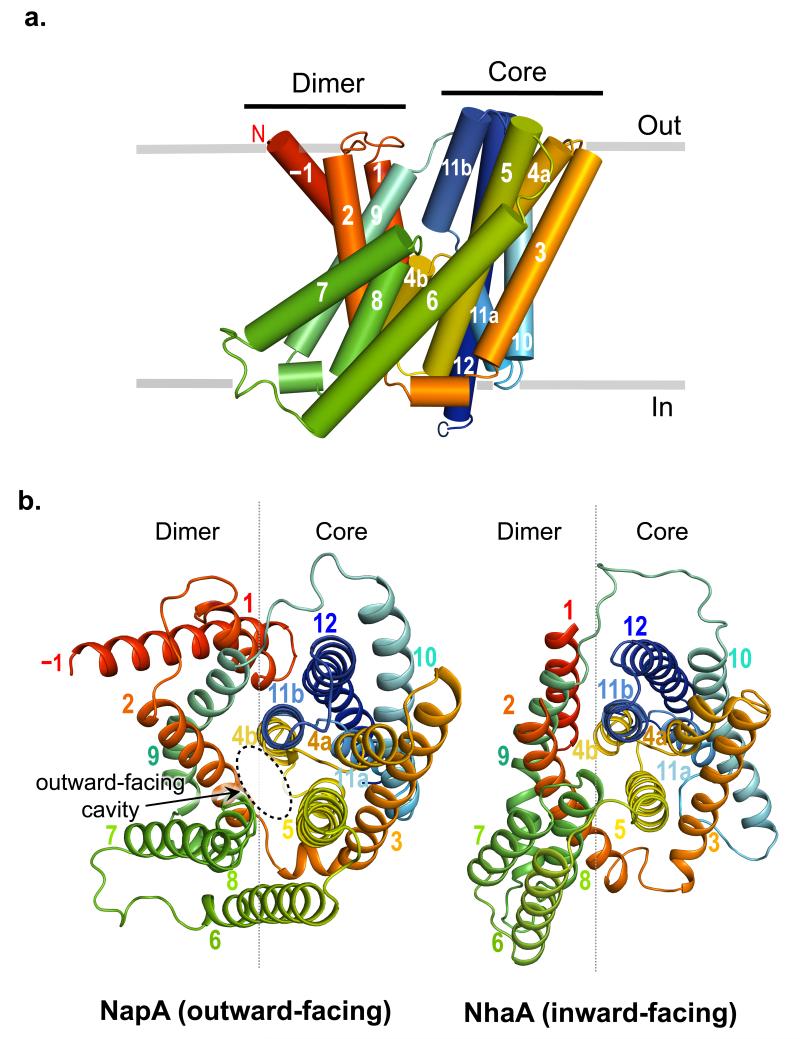
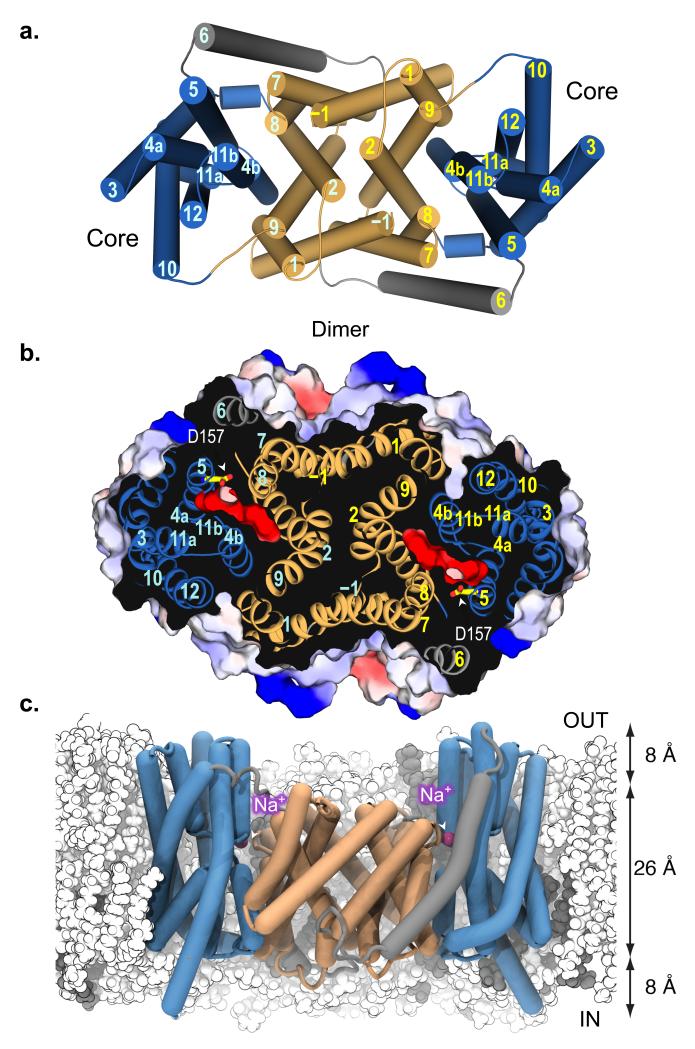
Sodium/proton (Na(+)/H(+)) antiporters, located at the plasma membrane in every cell, are vital for cell homeostasis. In humans, their dysfunction has been linked to diseases, such as hypertension, heart failure and epilepsy, and they are well-established drug targets. The best understood model system for Na(+)/H(+) antiport is NhaA from Escherichia coli, for which both electron microscopy and crystal structures are available. NhaA is made up of two distinct domains: a core domain and a dimerization domain. In the NhaA crystal structure a cavity is located between the two domains, providing access to the ion-binding site from the inward-facing surface of the protein. Like many Na(+)/H(+) antiporters, the activity of NhaA is regulated by pH, only becoming active above pH 6.5, at which point a conformational change is thought to occur. The only reported NhaA crystal structure so far is of the low pH inactivated form. Here we describe the active-state structure of a Na(+)/H(+) antiporter, NapA from Thermus thermophilus, at 3 Å resolution, solved from crystals grown at pH 7.8. In the NapA structure, the core and dimerization domains are in different positions to those seen in NhaA, and a negatively charged cavity has now opened to the outside. The extracellular cavity allows access to a strictly conserved aspartate residue thought to coordinate ion binding directly, a role supported here by molecular dynamics simulations. To alternate access to this ion-binding site, however, requires a surprisingly large rotation of the core domain, some 20° against the dimerization interface. We conclude that despite their fast transport rates of up to 1,500 ions per second, Na(+)/H(+) antiporters operate by a two-domain rocking bundle model, revealing themes relevant to secondary-active transporters in general.
钠离子/质子(Na(+)/H(+))反向转运体位于每个细胞的质膜上,对于细胞内稳态至关重要。在人类中,它们的功能障碍与高血压、心力衰竭和癫痫等疾病有关,它们是成熟的药物靶点。Na(+)/H(+)反向转运的最佳理解模型系统是来自大肠杆菌的 NhaA,它既有电子显微镜结构,也有晶体结构。NhaA 由两个不同的结构域组成:核心结构域和二聚化结构域。在 NhaA 的晶体结构中,两个结构域之间有一个空腔,从蛋白质的内向表面提供了进入离子结合位点的通道。与许多 Na(+)/H(+)反向转运体一样,NhaA 的活性受到 pH 的调节,只有在 pH 值高于 6.5 时才具有活性,此时被认为会发生构象变化。迄今为止,唯一报道的 NhaA 晶体结构是低 pH 失活形式。在这里,我们描述了来自嗜热栖热菌的 Na(+)/H(+)反向转运体 NapA 的活性状态结构,分辨率为 3Å,是在 pH 值为 7.8 时从晶体中解析出来的。在 NapA 结构中,核心和二聚化结构域的位置与 NhaA 不同,现在带负电荷的空腔已经向外部打开。细胞外空腔允许严格保守的天冬氨酸残基进入,该残基被认为直接协调离子结合,这里的分子动力学模拟支持了这一作用。然而,要交替进入这个离子结合位点,需要核心结构域惊人地旋转约 20°,与二聚化界面相对。我们得出的结论是,尽管 Na(+)/H(+)反向转运体的转运速度高达每秒 1500 个离子,但它们通过双域摇摆束模型起作用,揭示了一般来说与次级主动转运体相关的主题。