D'Alessio Cecilia, Dahms Nancy M
Laboratory of Glycobiology, Fundación Instituto Leloir - Instituto de Investigaciones Bioquimicas de Buenos Aires-CONICET, Av. Patricias Argentinas 435, C1405BWE, Buenos Aires, Argentina, and School of Sciences, University of Buenos Aires, C1428EHA, Buenos Aires, Argentina.
Curr Protein Pept Sci. 2015;16(1):31-48. doi: 10.2174/1389203716666150213160438.
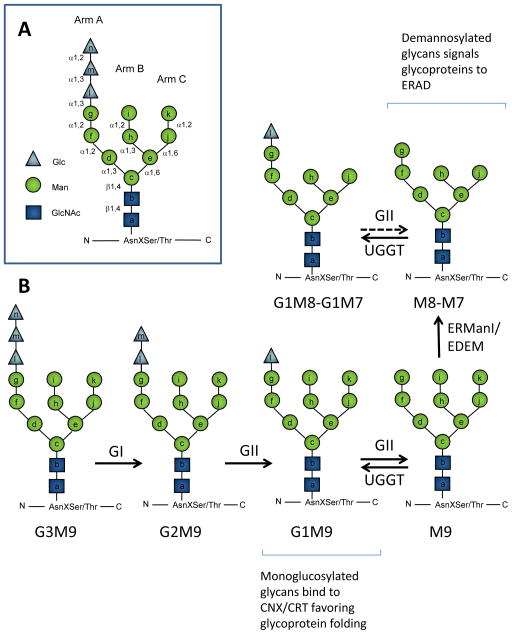
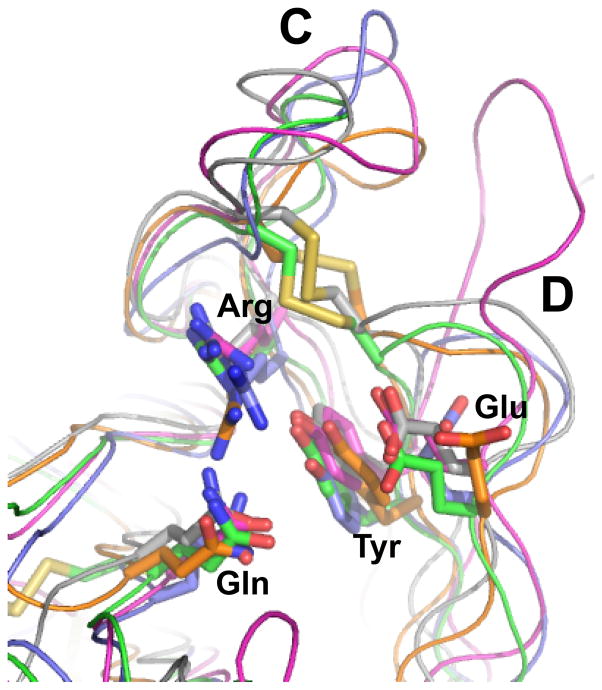
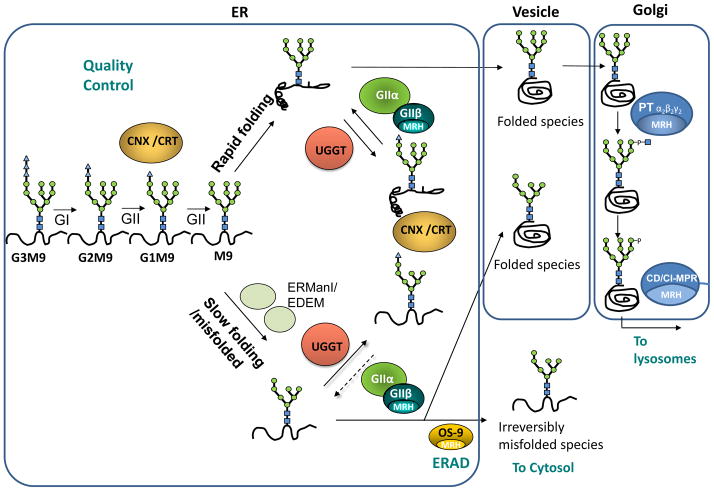
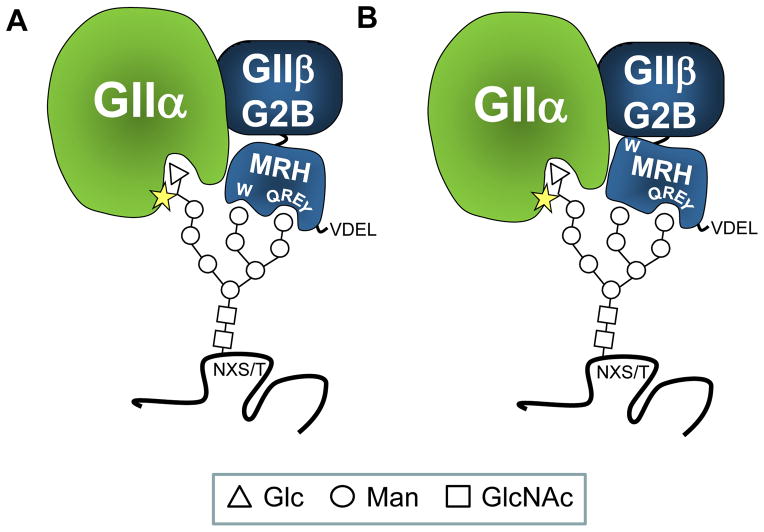
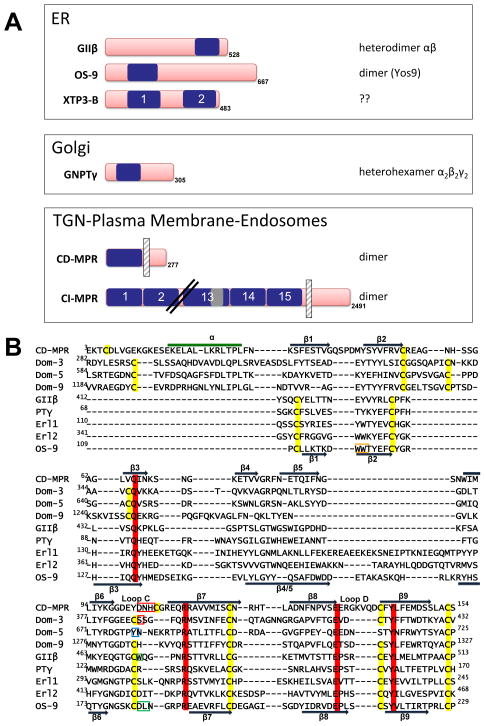
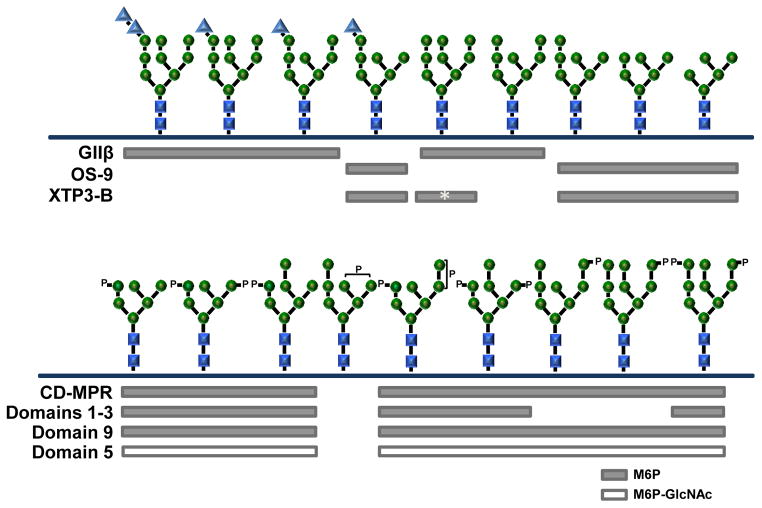
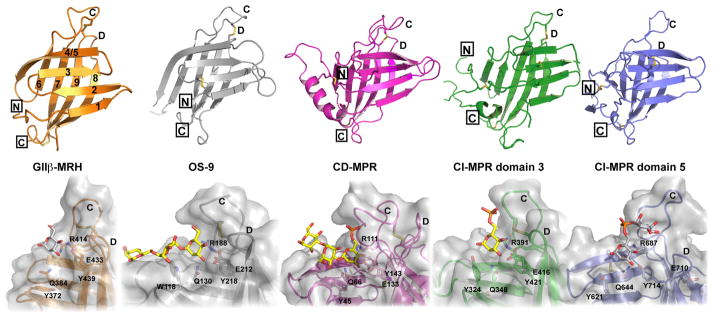
N-glycosylation in the endoplasmic reticulum (ER) consists of the transfer of a preassembled glycan conserved among species (Glc3Man9GlcNAc2) from a lipid donor to a consensus sequence within a nascent protein that is entering the ER. The protein-linked glycans are then processed by glycosidases and glycosyltransferases in the ER producing specific structures that serve as signalling molecules for the fate of the folding glycoprotein: to stay in the ER during the folding process, to be retrotranslocated to the cytosol for proteasomal degradation if irreversibly misfolded, or to pursue transit through the secretory pathway as a mature glycoprotein. In the ER, each glycan signalling structure is recognized by a specific lectin. A domain similar to that of the mannose 6-phosphate receptors (MPRs) has been identified in several proteins of the secretory pathway. These include the beta subunit of glucosidase II (GII), a key enzyme in the early processing of the transferred glycan that removes middle and innermost glucoses and is involved in quality control of glycoprotein folding in the ER (QC), the lectins OS-9 and XTP3-B, proteins involved in the delivery of ER misfolded proteins to degradation (ERAD), the gamma subunit of the Golgi GlcNAc-1-phosphotransferase, an enzyme involved in generating the mannose 6-phosphate (M6P) signal for sorting acidic hydrolases to lysosomes, and finally the MPRs that deliver those hydrolytic enzymes to the lysosome. Each of the MRH-containing proteins recognizes a different signalling N-glycan structure. Three-dimensional structures of some of the MRH domains have been solved, providing the basis to understand recognition mechanisms.
内质网(ER)中的N-糖基化包括将一种在物种间保守的预组装聚糖(Glc3Man9GlcNAc2)从脂质供体转移到进入内质网的新生蛋白质内的共有序列。然后,蛋白质连接的聚糖在内质网中由糖苷酶和糖基转移酶进行加工,产生特定结构,这些结构作为折叠糖蛋白命运的信号分子:在折叠过程中保留在内质网中,如果不可逆地错误折叠则逆向转运到细胞质中进行蛋白酶体降解,或者作为成熟糖蛋白通过分泌途径转运。在内质网中,每种聚糖信号结构都由一种特定的凝集素识别。在分泌途径的几种蛋白质中已鉴定出与甘露糖6-磷酸受体(MPR)结构域相似的结构域。这些包括葡糖苷酶II(GII)的β亚基,它是转移聚糖早期加工中的关键酶,可去除中间和最内层的葡萄糖,并参与内质网中糖蛋白折叠的质量控制(QC);凝集素OS-9和XTP3-B,参与将内质网错误折叠的蛋白质递送至降解(ERAD)的蛋白质;高尔基体GlcNAc-1-磷酸转移酶的γ亚基,一种参与生成将酸性水解酶分选到溶酶体的甘露糖6-磷酸(M6P)信号的酶;最后是将那些水解酶递送至溶酶体的MPR。每种含MRH的蛋白质都识别不同的信号N-聚糖结构。一些MRH结构域的三维结构已得到解析,为理解识别机制提供了基础。