Zheng Wang, Hussein Shaimaa, Yang JungWoo, Huang Jun, Zhang Fan, Hernandez-Anzaldo Samuel, Fernandez-Patron Carlos, Cao Ying, Zeng Hongbo, Tang Jingfeng, Chen Xing-Zhen
Membrane Protein Disease Research Group, Department of Physiology, Faculty of Medicine and Dentistry, University of Alberta, T6G 2H7, Edmonton, AB, Canada.
Department of Chemical and Materials Engineering, Faculty of Engineering, University of Alberta, T6G 2V4, Edmonton, AB, Canada.
Sci Rep. 2015 Mar 30;5:9460. doi: 10.1038/srep09460.
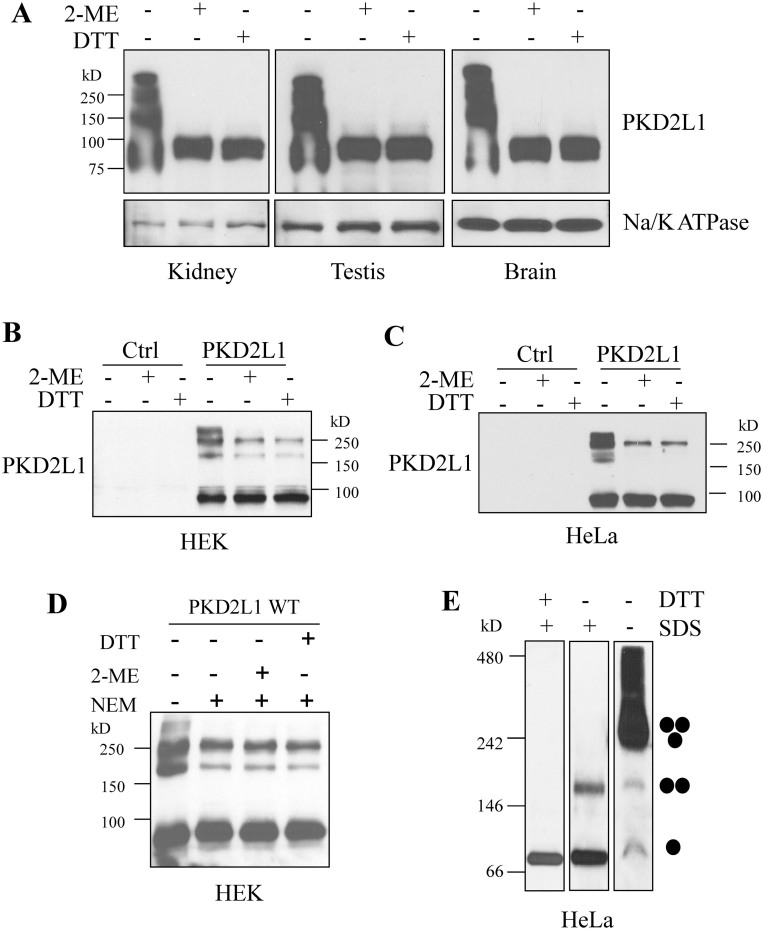
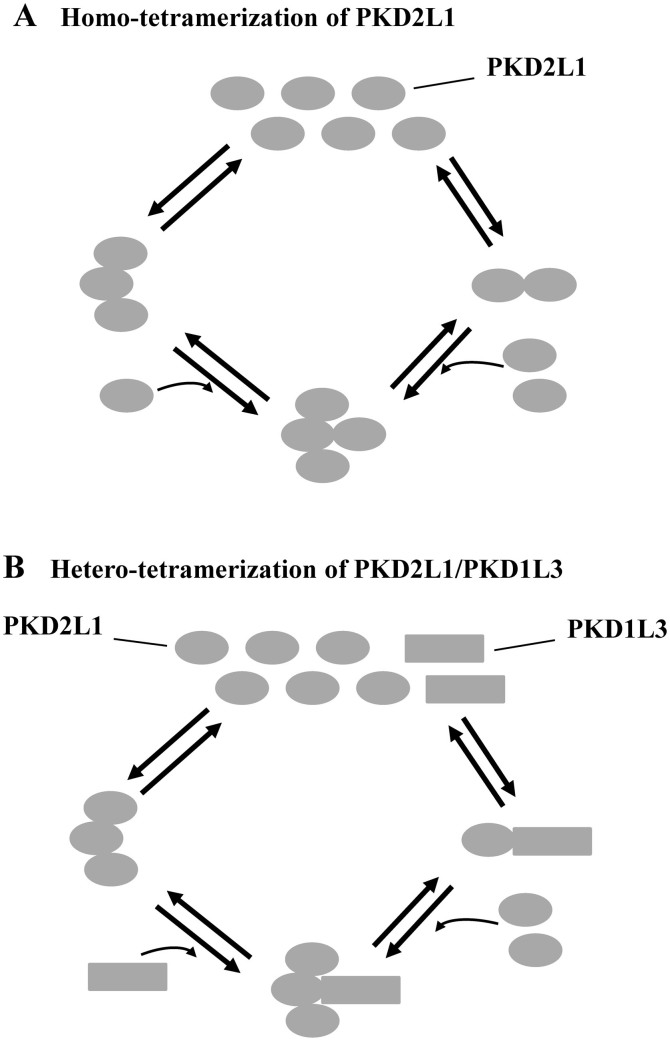
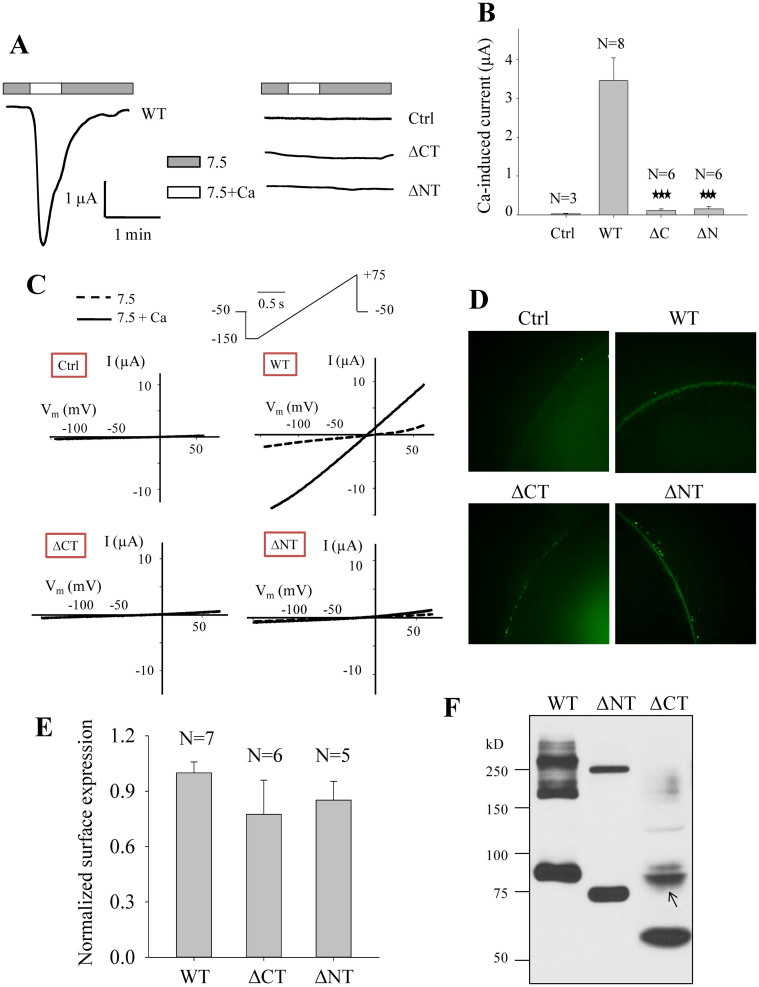
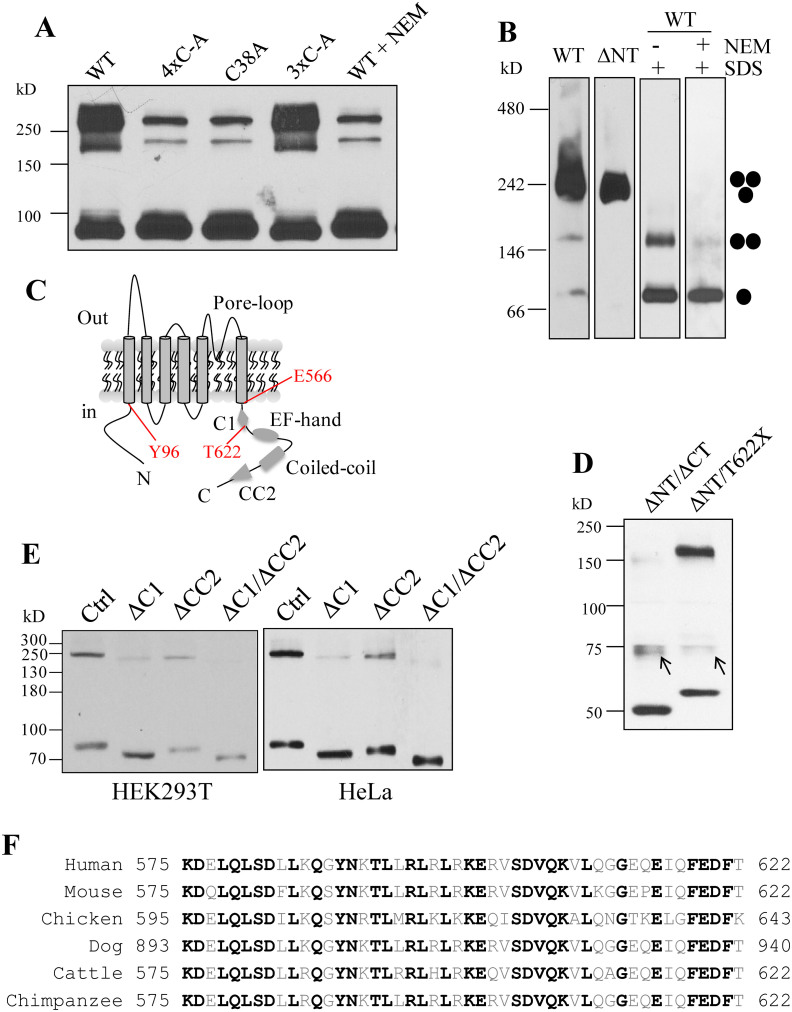
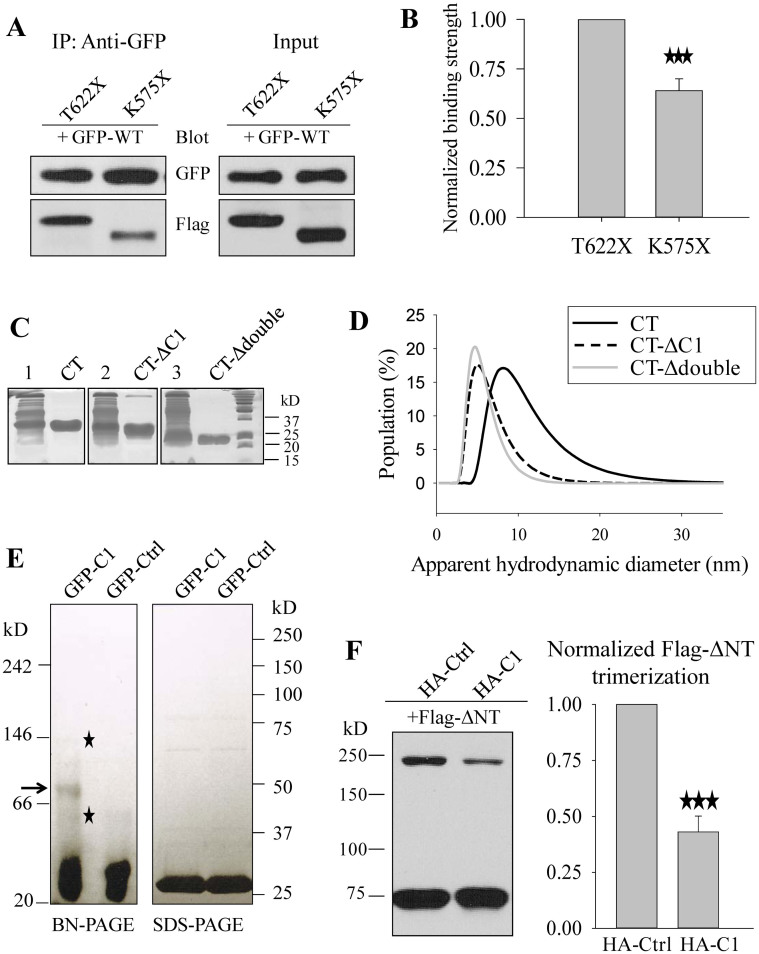
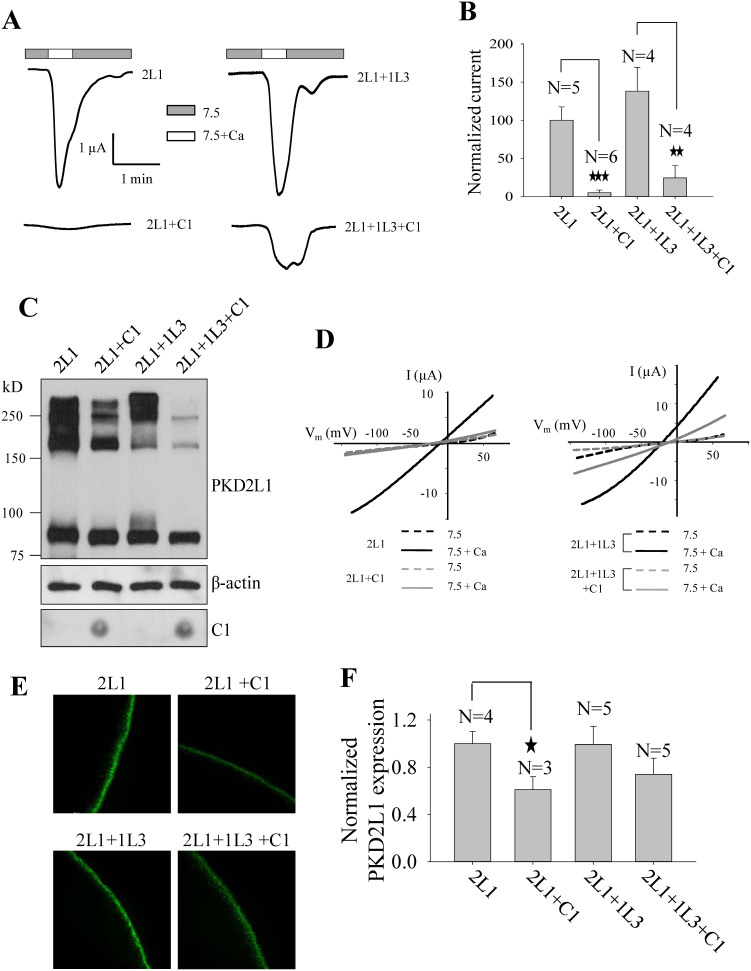
As a transient receptor potential (TRP) superfamily member, polycystic kidney disease 2-like-1 (PKD2L1) is also called TRPP3 and has similar membrane topology as voltage-gated cation channels. PKD2L1 is involved in hedgehog signaling, intestinal development, and sour tasting. PKD2L1 and PKD1L3 form heterotetramers with 3:1 stoichiometry. C-terminal coiled-coil-2 (CC2) domain (G699-W743) of PKD2L1 was reported to be important for its trimerization but independent studies showed that CC2 does not affect PKD2L1 channel function. It thus remains unclear how PKD2L1 proteins oligomerize into a functional channel. By SDS-PAGE, blue native PAGE and mutagenesis we here identified a novel C-terminal domain called C1 (K575-T622) involved in stronger homotrimerization than the non-overlapping CC2, and found that the PKD2L1 N-terminus is critical for dimerization. By electrophysiology and Xenopus oocyte expression, we found that C1, but not CC2, is critical for PKD2L1 channel function. Our co-immunoprecipitation and dynamic light scattering experiments further supported involvement of C1 in trimerization. Further, C1 acted as a blocking peptide that inhibits PKD2L1 trimerization as well as PKD2L1 and PKD2L1/PKD1L3 channel function. Thus, our study identified C1 as the first PKD2L1 domain essential for both PKD2L1 trimerization and channel function, and suggest that PKD2L1 and PKD2L1/PKD1L3 channels share the PKD2L1 trimerization process.
作为瞬时受体电位(TRP)超家族成员,多囊肾病2样-1(PKD2L1)也被称为TRPP3,具有与电压门控阳离子通道相似的膜拓扑结构。PKD2L1参与刺猬信号通路、肠道发育和酸味感知。PKD2L1和PKD1L3以3:1的化学计量比形成异源四聚体。据报道,PKD2L1的C末端卷曲螺旋-2(CC2)结构域(G699-W743)对其三聚化很重要,但独立研究表明CC2不影响PKD2L1通道功能。因此,PKD2L1蛋白如何寡聚形成功能性通道仍不清楚。通过SDS-PAGE、蓝色天然PAGE和诱变,我们在此鉴定了一个新的C末端结构域,称为C1(K575-T622),它比不重叠的CC2参与更强的同源三聚化,并发现PKD2L1的N末端对二聚化至关重要。通过电生理学和非洲爪蟾卵母细胞表达,我们发现C1而非CC2对PKD2L1通道功能至关重要。我们的免疫共沉淀和动态光散射实验进一步支持了C1参与三聚化。此外,C1作为一种阻断肽,可抑制PKD2L1三聚化以及PKD2L1和PKD2L1/PKD1L3通道功能。因此,我们的研究确定C1是PKD2L1三聚化和通道功能所必需的首个PKD2L1结构域,并表明PKD2L1和PKD2L1/PKD1L3通道共享PKD2L1三聚化过程。