Fanti Federico, Cau Andrea, Cantelli Luigi, Hassine Mohsen, Auditore Marco
Dipartimento di Scienze Biologiche, Geologiche e Ambientali, Alma Mater Studiorum, Università di Bologna, Bologna, Italy; Museo Geologico Giovanni Capellini, Alma Mater Studiorum, Università di Bologna, Bologna, Italy.
Dipartimento di Scienze Biologiche, Geologiche e Ambientali, Alma Mater Studiorum, Università di Bologna, Bologna, Italy.
PLoS One. 2015 Apr 29;10(4):e0123475. doi: 10.1371/journal.pone.0123475. eCollection 2015.
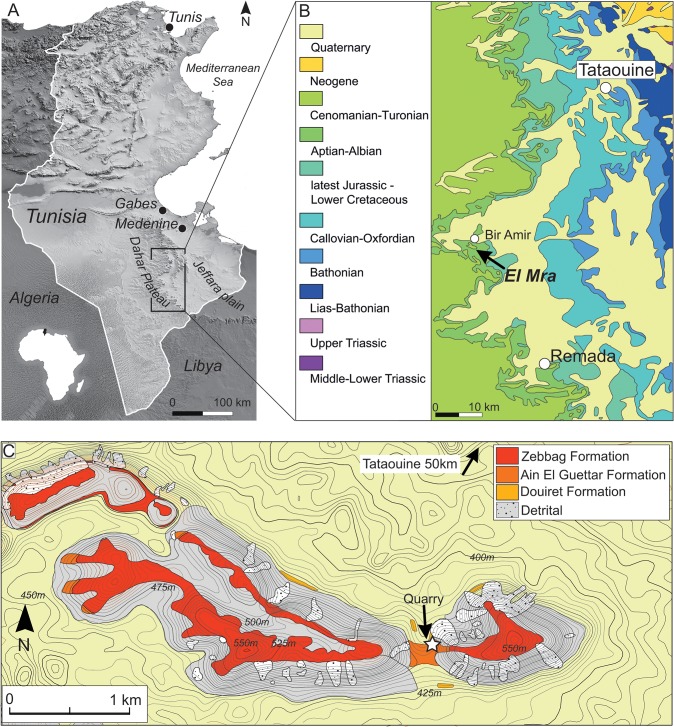
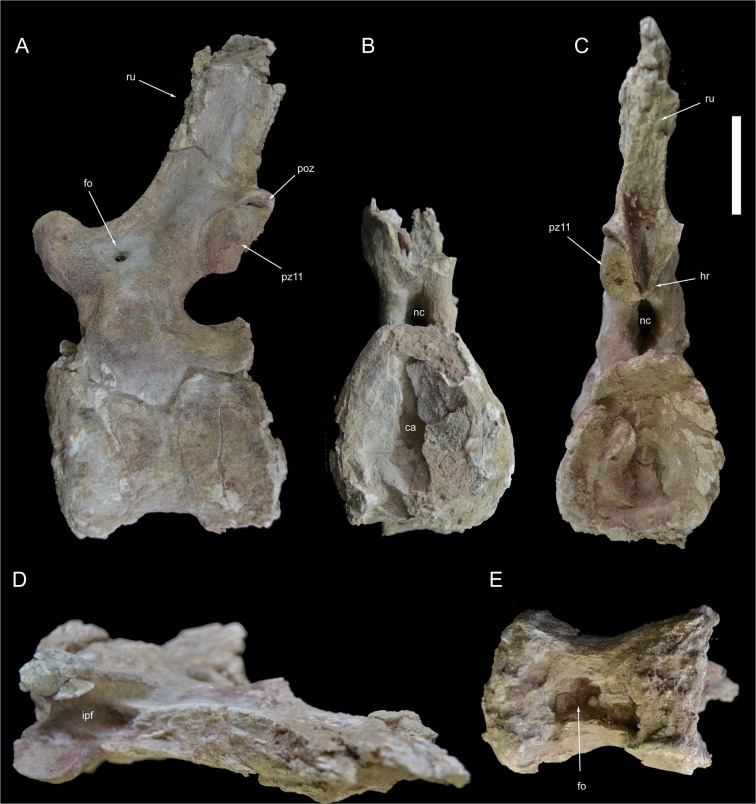
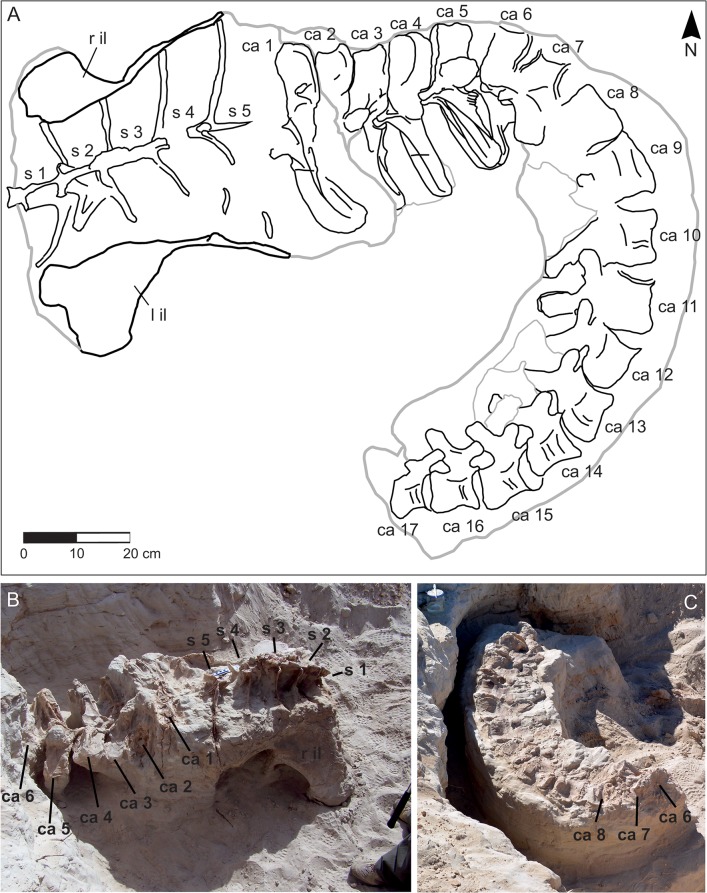
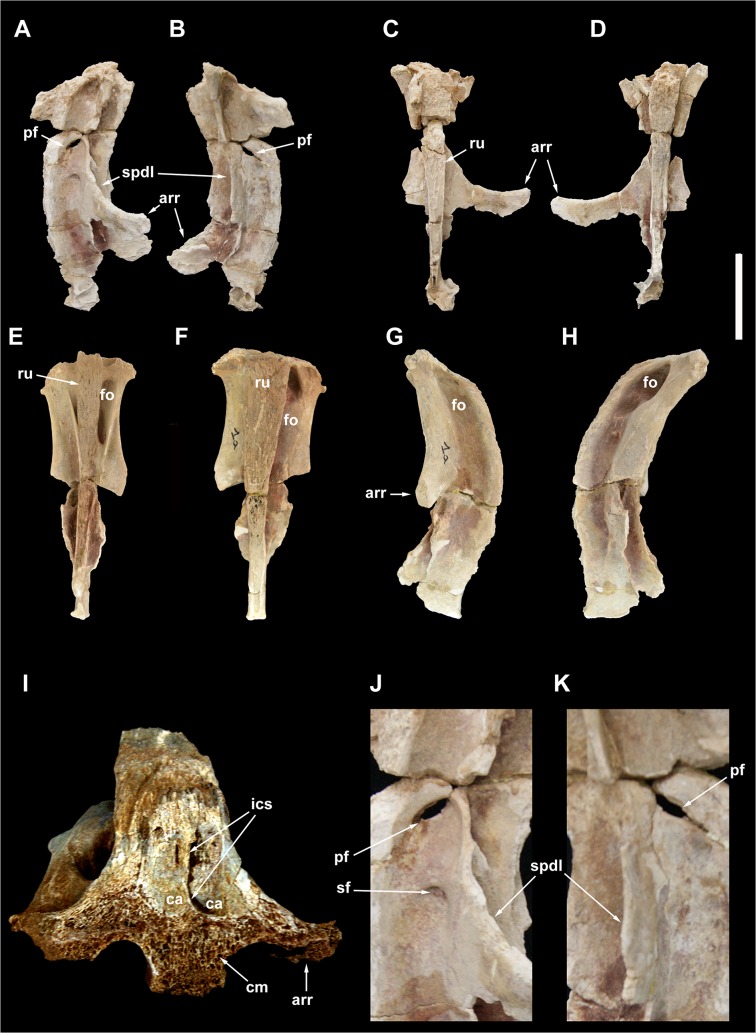
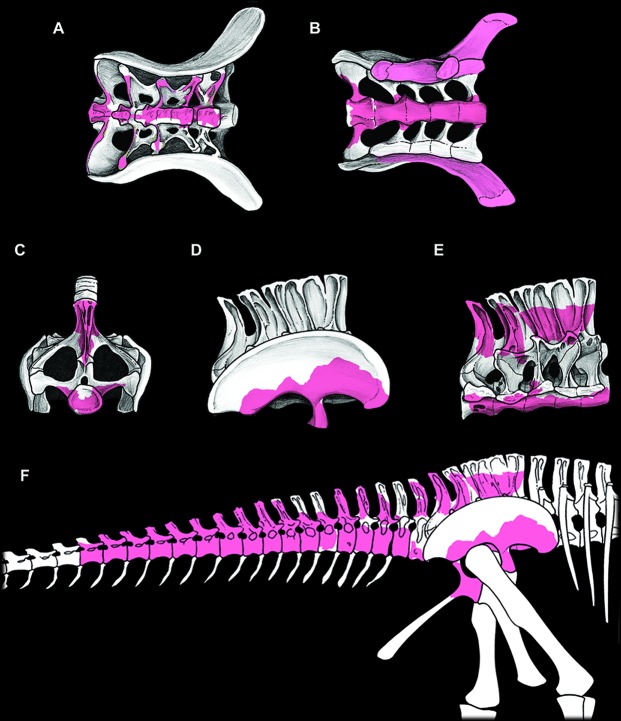
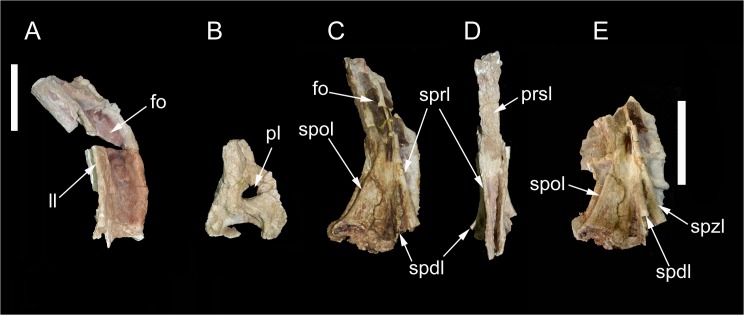
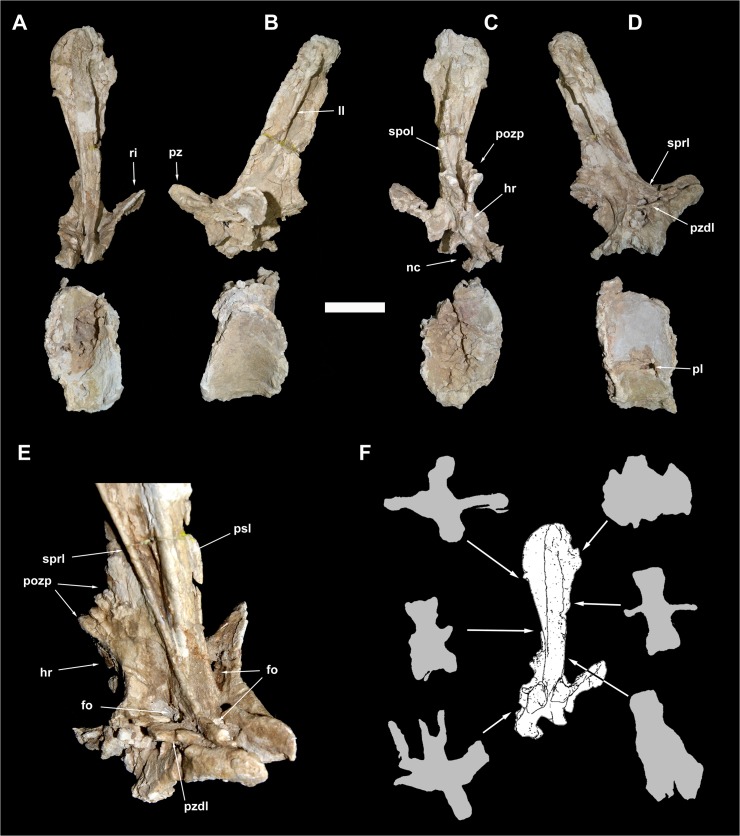
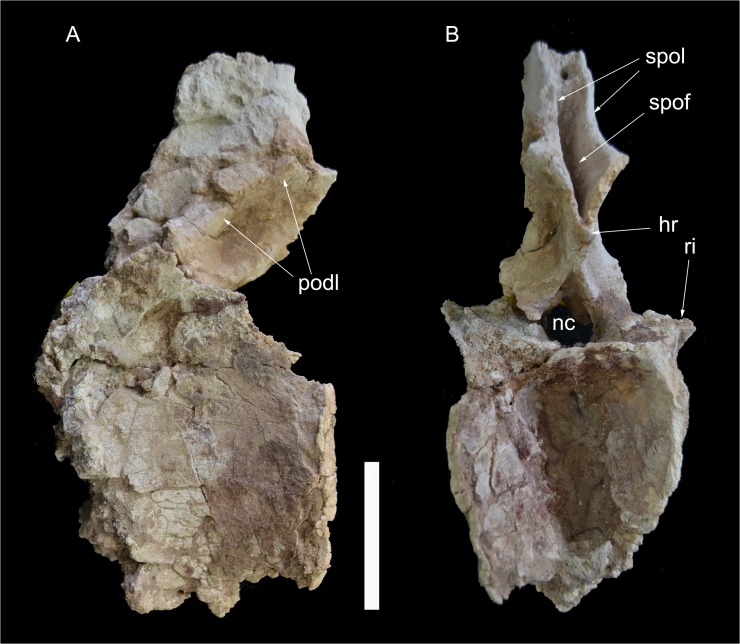
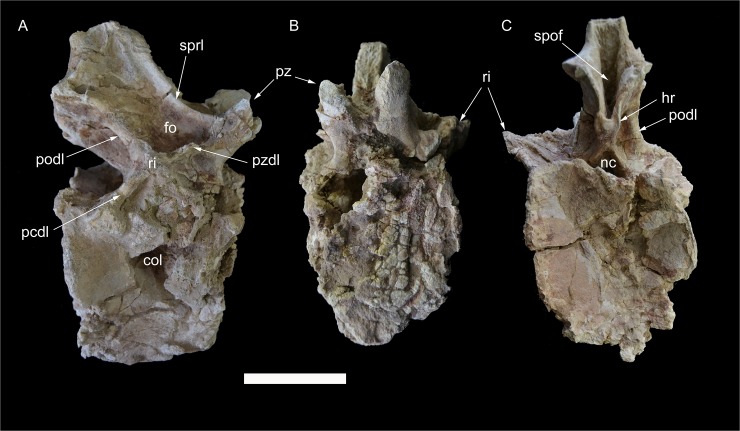
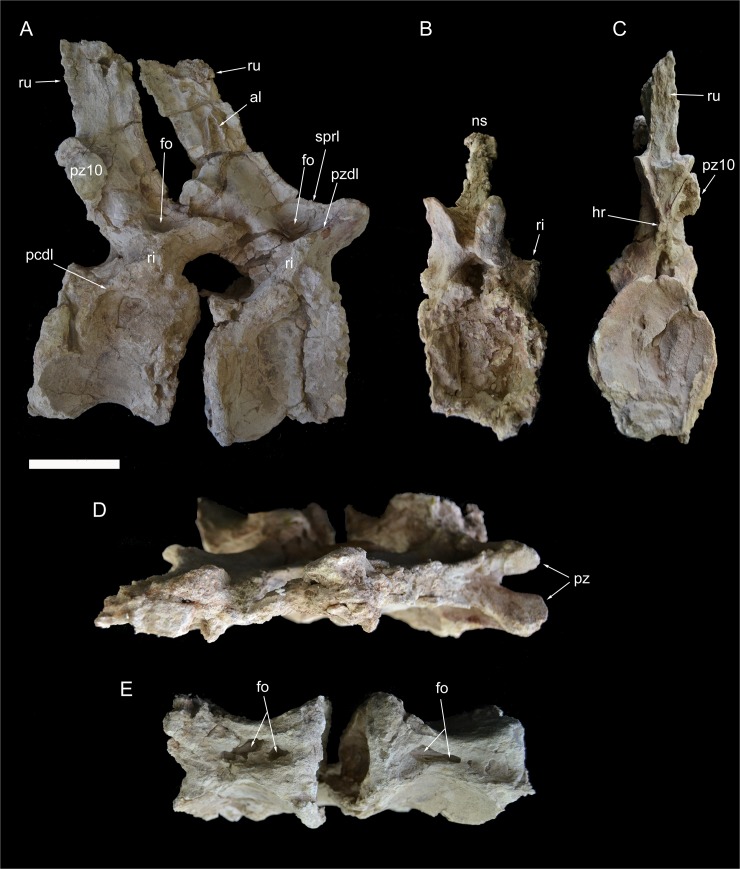
The rebbachisaurid sauropod Tataouinea hannibalis represents the first articulated dinosaur skeleton from Tunisia and one of the best preserved in northern Africa. The type specimen was collected from the lower Albian, fluvio-estuarine deposits of the Ain el Guettar Formation (southern Tunisia). We present detailed analyses on the sedimentology and facies distribution at the main quarry and a revision of the vertebrate fauna associated with the skeleton. Data provide information on a complex ecosystem dominated by crocodilian and other brackish water taxa. Taphonomic interpretations indicate a multi-event, pre-burial history with a combination of rapid segregation in high sediment supply conditions and partial subaerial exposure of the carcass. After the collection in 2011 of the articulated sacrum and proximalmost caudal vertebrae, all showing a complex pattern of pneumatization, newly discovered material of the type specimen allows a detailed osteological description of Tataouinea. The sacrum, the complete and articulated caudal vertebrae 1-17, both ilia and ischia display asymmetrical pneumatization, with the left side of vertebrae and the left ischium showing a more extensive invasion by pneumatic features than their right counterparts. A pneumatic hiatus is present in caudal centra 7 to 13, whereas caudal centra 14-16 are pneumatised by shallow fossae. Bayesian inference analyses integrating morphological, stratigraphic and paleogeographic data support a flagellicaudatan-rebbachisaurid divergence at about 163 Ma and a South American ancestral range for rebbachisaurids. Results presented here suggest an exclusively South American Limaysaurinae and a more widely distributed Rebbachisaurinae lineage, the latter including the South American taxon Katepensaurus and a clade including African and European taxa, with Tataouinea as sister taxon of Rebbachisaurus. This scenario would indicate that South America was not affected by the end-Jurassic extinction of diplodocoids, and was most likely the centre of the rapid radiation of rebbachisaurids to Africa and Europe between 135 and 130 Ma.
雷巴齐斯龙科的蜥脚类恐龙汉尼拔塔陶内龙是来自突尼斯的首个连接完整的恐龙骨架,也是北非保存最完好的恐龙骨架之一。模式标本采自艾因盖塔尔组(突尼斯南部)下阿尔比阶的河海相沉积层。我们对主采石场的沉积学和相分布进行了详细分析,并对与该骨架相关的脊椎动物群进行了修订。数据提供了一个以鳄鱼和其他微咸水类群为主导的复杂生态系统的信息。埋藏学解释表明,该恐龙尸体在被掩埋之前经历了多起事件,先是在沉积物供应量大的情况下迅速分离,之后尸体部分暴露于空气中。2011年采集到连接完整的骶骨和最前端的尾椎骨,所有这些骨骼都呈现出复杂的气腔化模式,新发现的模式标本材料使得对汉尼拔塔陶内龙进行详细的骨骼学描述成为可能。骶骨、完整且连接在一起的第1至17节尾椎骨、双侧髂骨和坐骨均显示出不对称的气腔化,椎体左侧和左侧坐骨的气腔特征比右侧更为广泛。第7至13节尾椎椎体存在气腔间隙,而第14至16节尾椎椎体则由浅窝气腔化。整合形态学、地层学和古地理数据的贝叶斯推断分析支持鞭尾类与雷巴齐斯龙科在约1.63亿年前分化,且雷巴齐斯龙科的祖先分布在南美洲。此处给出的结果表明,利迈龙亚科仅分布于南美洲,而雷巴齐斯龙亚科的分布更为广泛,后者包括南美洲的分类单元卡特彭龙以及一个包含非洲和欧洲分类单元的分支,汉尼拔塔陶内龙是雷巴齐斯龙的姐妹分类单元。这种情况表明,南美洲未受到侏罗纪末期梁龙类灭绝事件的影响,并且很可能是1.35亿至1.30亿年前雷巴齐斯龙科迅速向非洲和欧洲辐射的中心。