McCarthy Elizabeth W, Arnold Sarah E J, Chittka Lars, Le Comber Steven C, Verity Robert, Dodsworth Steven, Knapp Sandra, Kelly Laura J, Chase Mark W, Baldwin Ian T, Kovařík Aleš, Mhiri Corinne, Taylor Lin, Leitch Andrew R
School of Biological and Chemical Sciences, Queen Mary University of London, Mile End Road, London E1 4NS, UK, Natural History Museum, London SW7 5BD, UK, Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3DS, UK, Max Planck Institute for Chemical Ecology, Department of Molecular Ecology, Beutenberg Campus, Hans-Knöll-Strasse 8, 07745 Jena, Germany, Institute of Biophysics, Academy of Sciences of the Czech Republic, CZ-61265 Brno, Czech Republic, Institut Jean-Pierre Bourgin, UMR1318 INRA-AgroParisTech, INRA-Versailles, 78026 Versailles cedex, France and Department of Plant Sciences, University of Cambridge, Downing Street, Cambridge CB2 3EA, UK School of Biological and Chemical Sciences, Queen Mary University of London, Mile End Road, London E1 4NS, UK, Natural History Museum, London SW7 5BD, UK, Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3DS, UK, Max Planck Institute for Chemical Ecology, Department of Molecular Ecology, Beutenberg Campus, Hans-Knöll-Strasse 8, 07745 Jena, Germany, Institute of Biophysics, Academy of Sciences of the Czech Republic, CZ-61265 Brno, Czech Republic, Institut Jean-Pierre Bourgin, UMR1318 INRA-AgroParisTech, INRA-Versailles, 78026 Versailles cedex, France and Department of Plant Sciences, University of Cambridge, Downing Street, Cambridge CB2 3EA, UK.
School of Biological and Chemical Sciences, Queen Mary University of London, Mile End Road, London E1 4NS, UK, Natural History Museum, London SW7 5BD, UK, Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3DS, UK, Max Planck Institute for Chemical Ecology, Department of Molecular Ecology, Beutenberg Campus, Hans-Knöll-Strasse 8, 07745 Jena, Germany, Institute of Biophysics, Academy of Sciences of the Czech Republic, CZ-61265 Brno, Czech Republic, Institut Jean-Pierre Bourgin, UMR1318 INRA-AgroParisTech, INRA-Versailles, 78026 Versailles cedex, France and Department of Plant Sciences, University of Cambridge, Downing Street, Cambridge CB2 3EA, UK.
Ann Bot. 2015 Jun;115(7):1117-31. doi: 10.1093/aob/mcv048. Epub 2015 May 15.
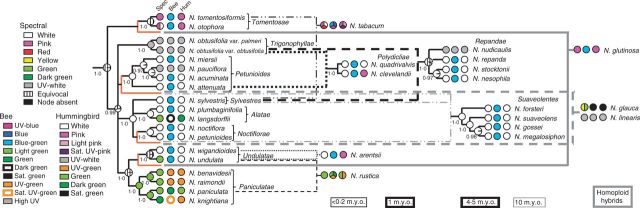
Speciation in angiosperms can be accompanied by changes in floral colour that may influence pollinator preference and reproductive isolation. This study investigates whether changes in floral colour can accompany polyploid and homoploid hybridization, important processes in angiosperm evolution.
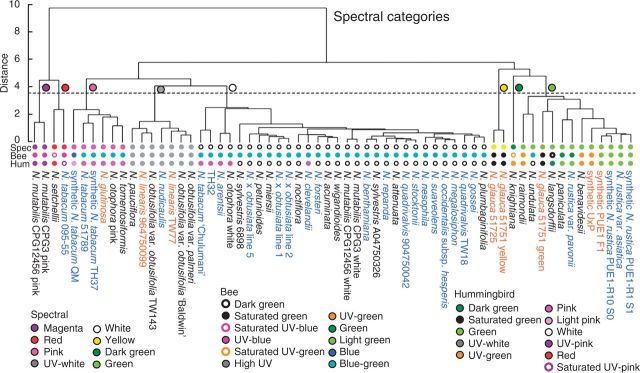
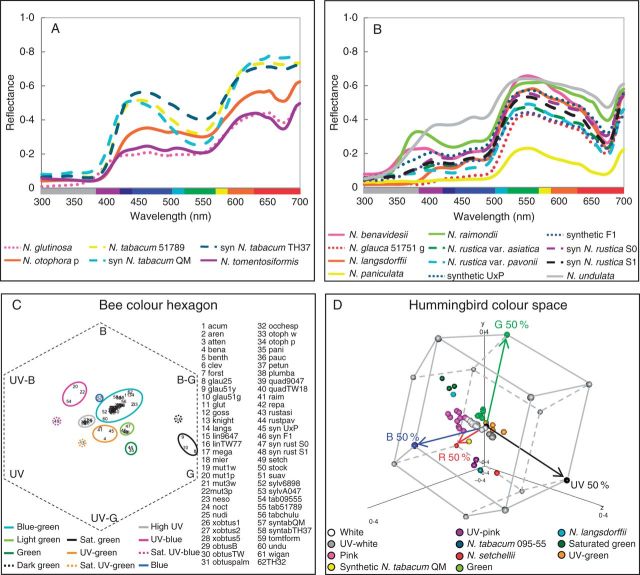
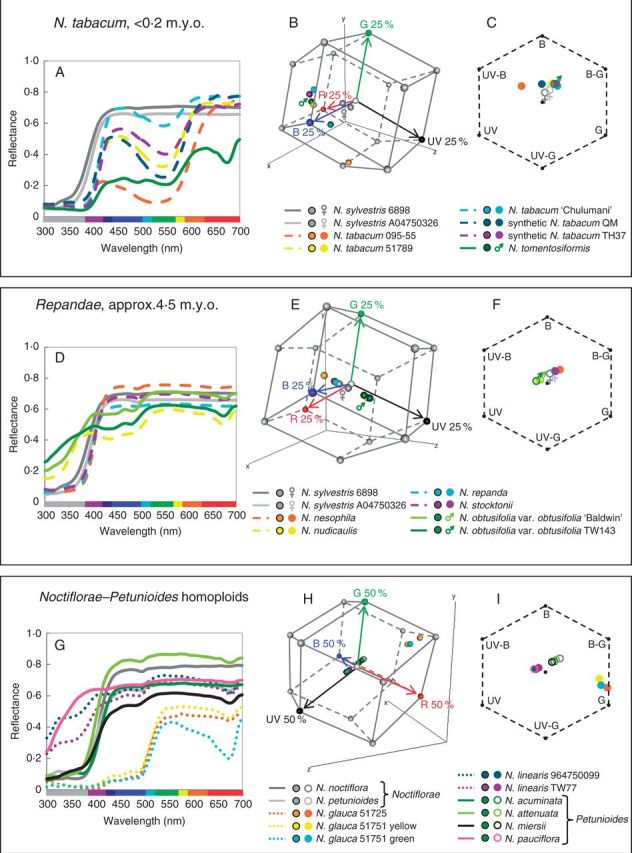
Spectral reflectance of corolla tissue was examined for 60 Nicotiana (Solanaceae) accessions (41 taxa) based on spectral shape (corresponding to pigmentation) as well as bee and hummingbird colour perception in order to assess patterns of floral colour evolution. Polyploid and homoploid hybrid spectra were compared with those of their progenitors to evaluate whether hybridization has resulted in floral colour shifts.
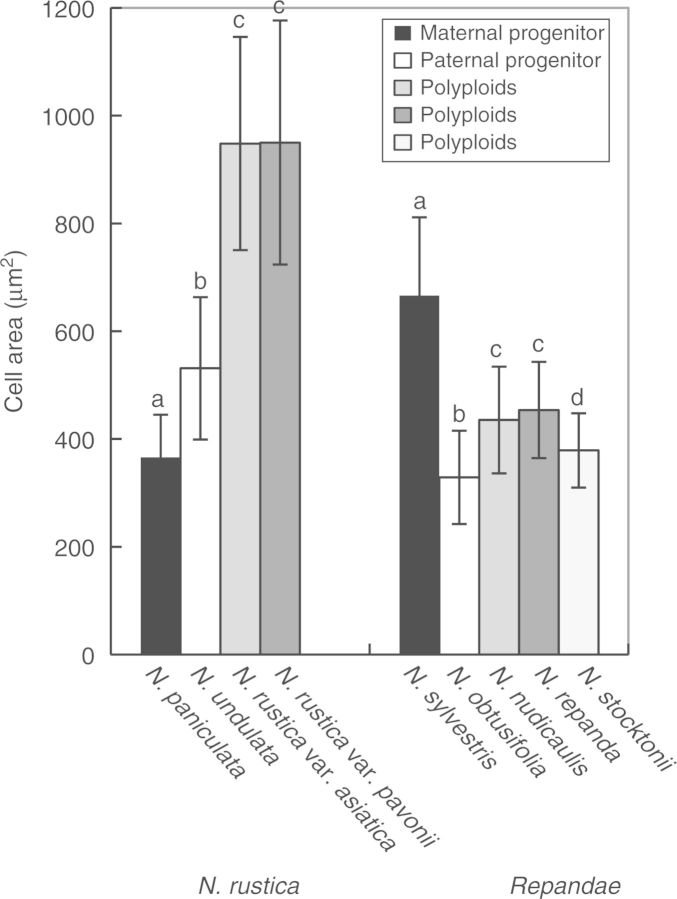
Floral colour categories in Nicotiana seem to have arisen multiple times independently during the evolution of the genus. Most younger polyploids displayed an unexpected floral colour, considering those of their progenitors, in the colour perception of at least one pollinator type, whereas older polyploids tended to resemble one or both of their progenitors.
Floral colour evolution in Nicotiana is weakly constrained by phylogeny, and colour shifts do occur in association with both polyploid and homoploid hybrid divergence. Transgressive floral colour in N. tabacum has arisen by inheritance of anthocyanin pigmentation from its paternal progenitor while having a plastid phenotype like its maternal progenitor. Potentially, floral colour evolution has been driven by, or resulted in, pollinator shifts. However, those polyploids that are not sympatric (on a regional scale) with their progenitor lineages are typically not divergent in floral colour from them, perhaps because of a lack of competition for pollinators.
被子植物的物种形成可能伴随着花色变化,这可能会影响传粉者偏好和生殖隔离。本研究调查花色变化是否会伴随多倍体和同倍体杂交发生,这是被子植物进化中的重要过程。
基于光谱形状(对应色素沉着)以及蜜蜂和蜂鸟的颜色感知,对60份烟草属(茄科)种质(41个分类群)的花冠组织光谱反射率进行了检测,以评估花色进化模式。将多倍体和同倍体杂种的光谱与其亲本的光谱进行比较,以评估杂交是否导致了花色变化。
在烟草属的进化过程中,花色类别似乎多次独立出现。考虑到其亲本的花色,大多数较年轻的多倍体在至少一种传粉者类型的颜色感知中呈现出意想不到的花色,而较古老的多倍体则倾向于与其一个或两个亲本相似。
烟草属的花色进化受系统发育的限制较弱,花色变化确实与多倍体和同倍体杂种分化有关。烟草的超亲花色是通过从其父本亲本继承花青素色素沉着而产生的,同时具有与其母本亲本相似的质体表型。花色进化可能是由传粉者转变驱动的,或者导致了传粉者转变。然而,那些在区域尺度上与其亲本谱系不同域的多倍体,其花色通常与其亲本没有差异,这可能是因为缺乏对传粉者的竞争。