Gil-Ranedo Jon, Hernando Eva, Riolobos Laura, Domínguez Carlos, Kann Michael, Almendral José M
Centro de Biología Molecular "Severo Ochoa" (Consejo Superior de Investigaciones Científicas-Universidad Autónoma de Madrid), Cantoblanco, Madrid, Spain.
University of Bordeaux, Microbiologie Fondamentale et Pathogénicité, UMR 5234, Bordeaux, France; CNRS, Microbiologie Fondamentale et Pathogénicité, UMR 5234, Bordeaux, France; Centre Hospitalier Universitaire de Bordeaux, Service de Virologie, Bordeaux, France.
PLoS Pathog. 2015 Jun 11;11(6):e1004920. doi: 10.1371/journal.ppat.1004920. eCollection 2015 Jun.
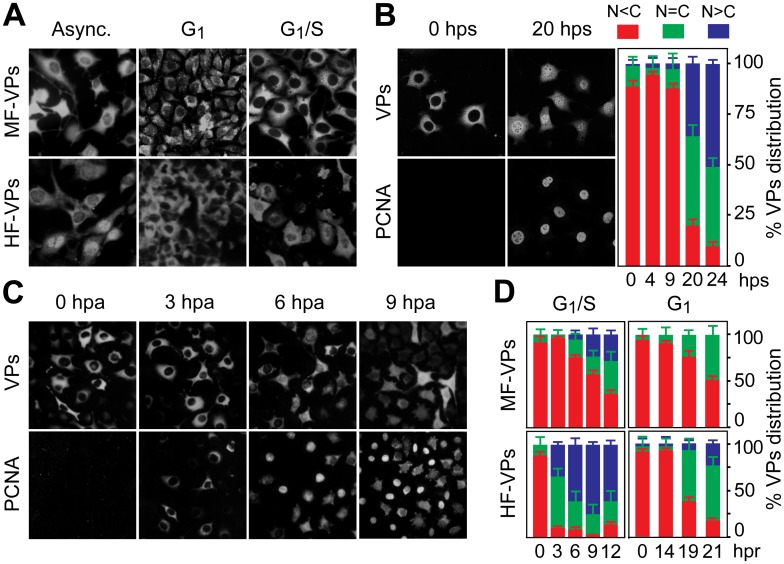
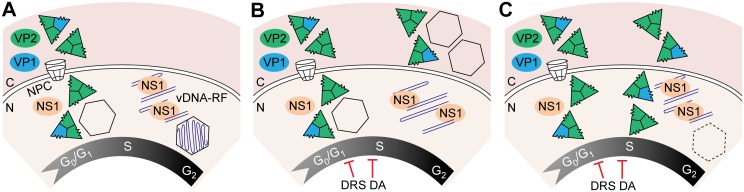
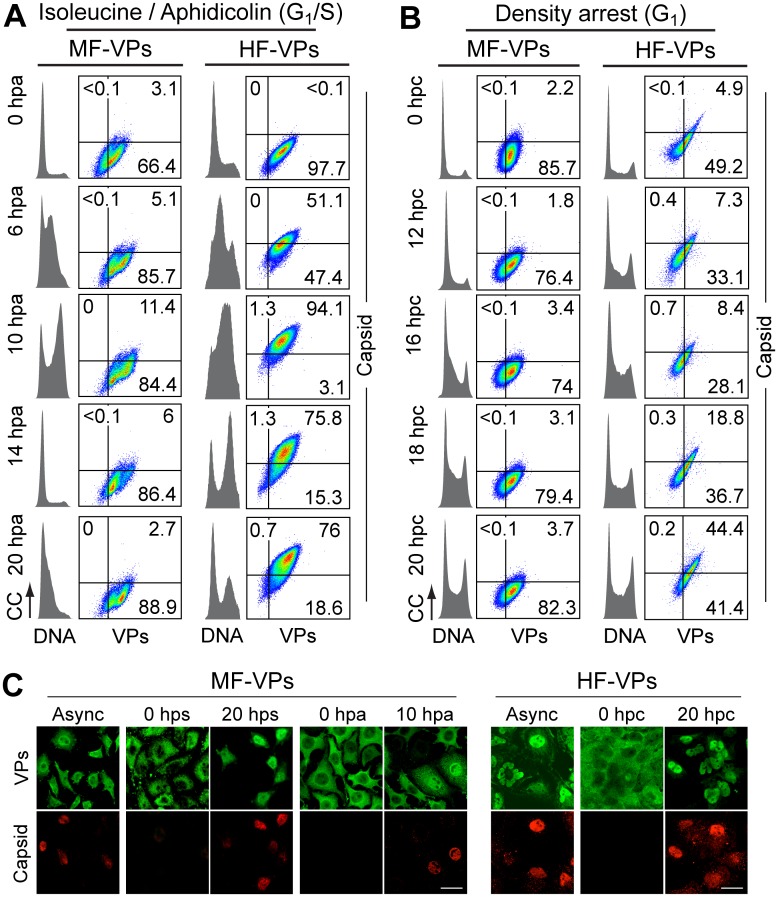
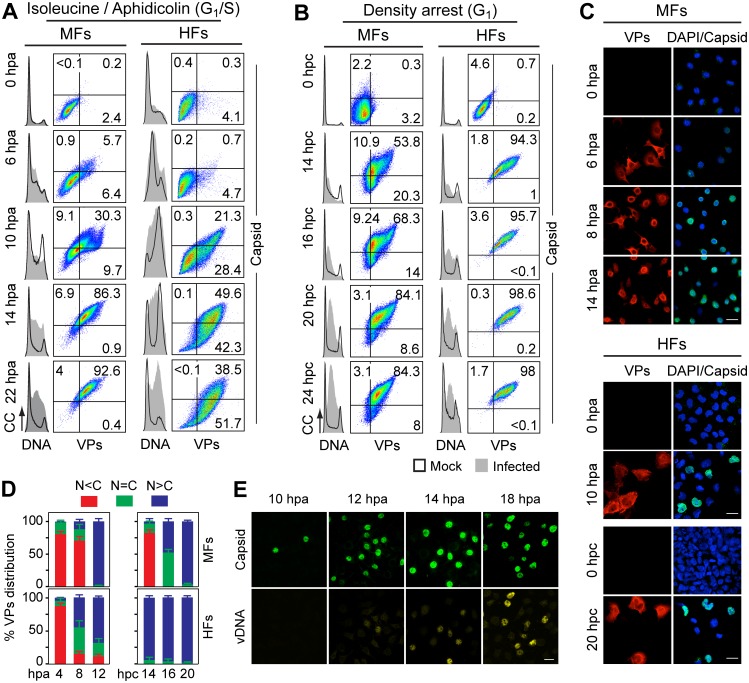
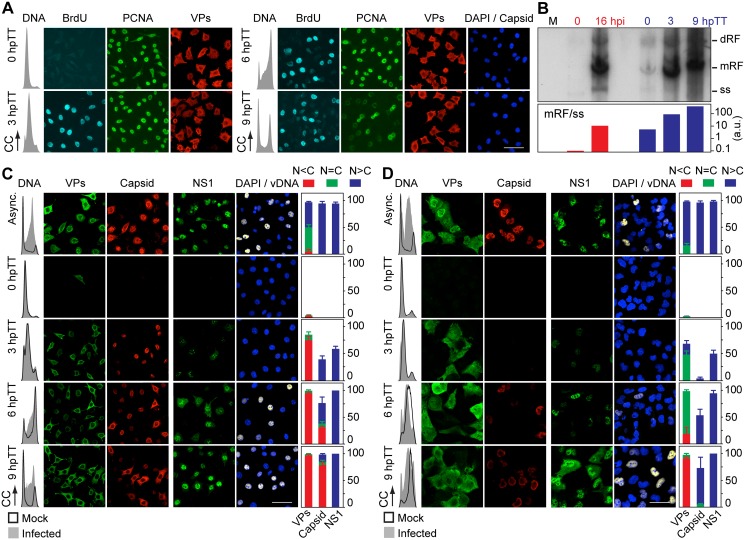
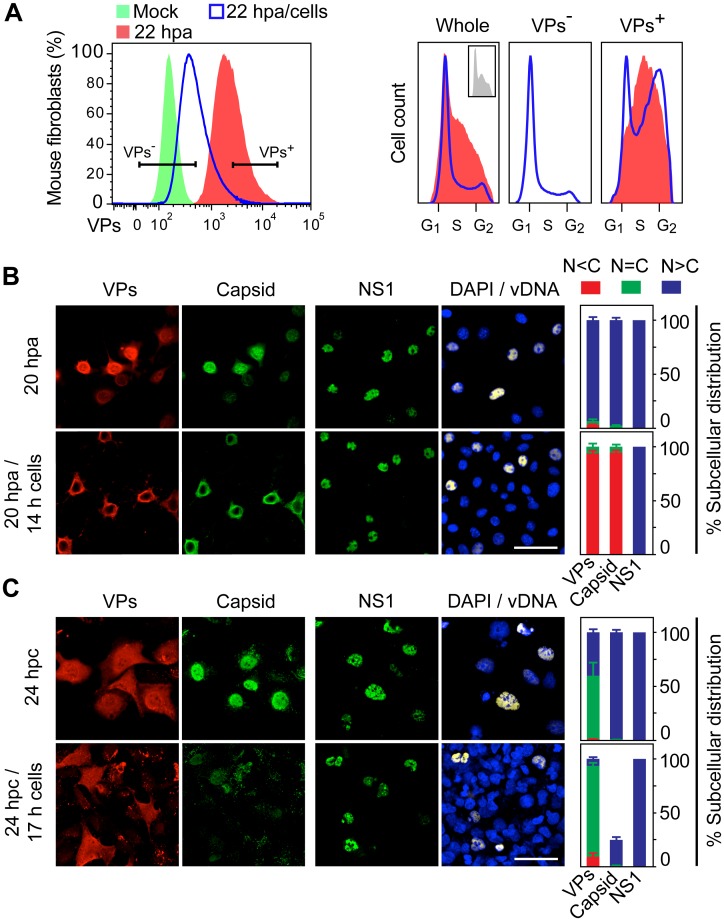
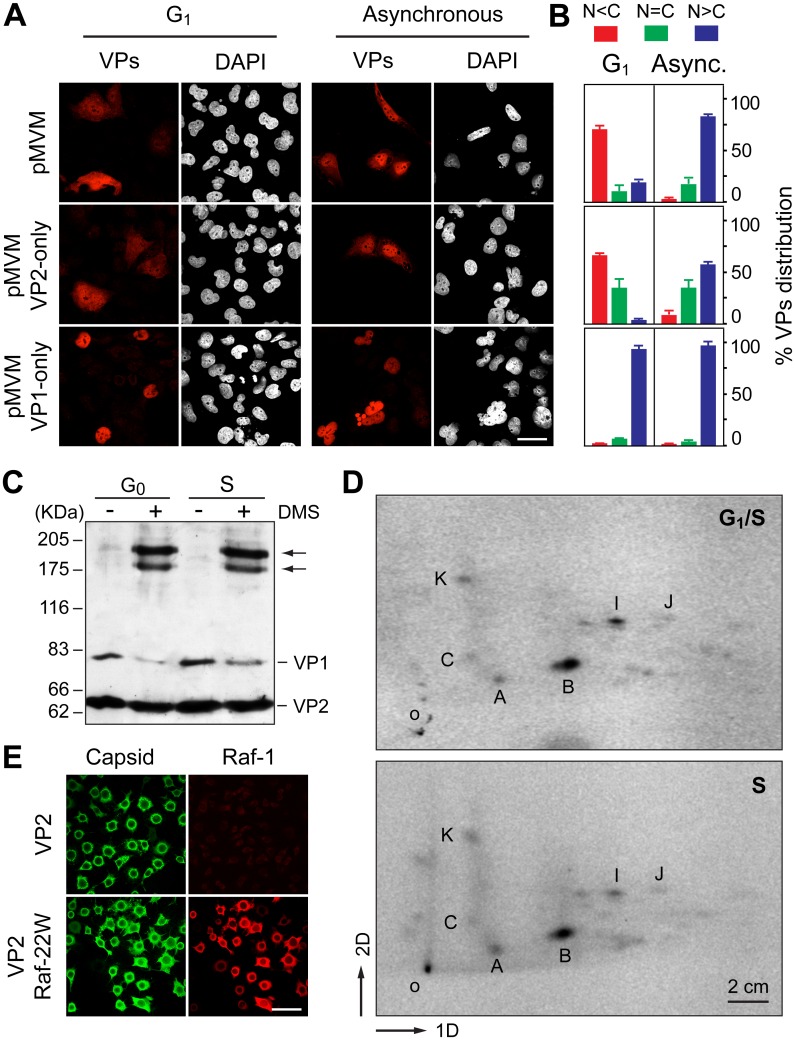
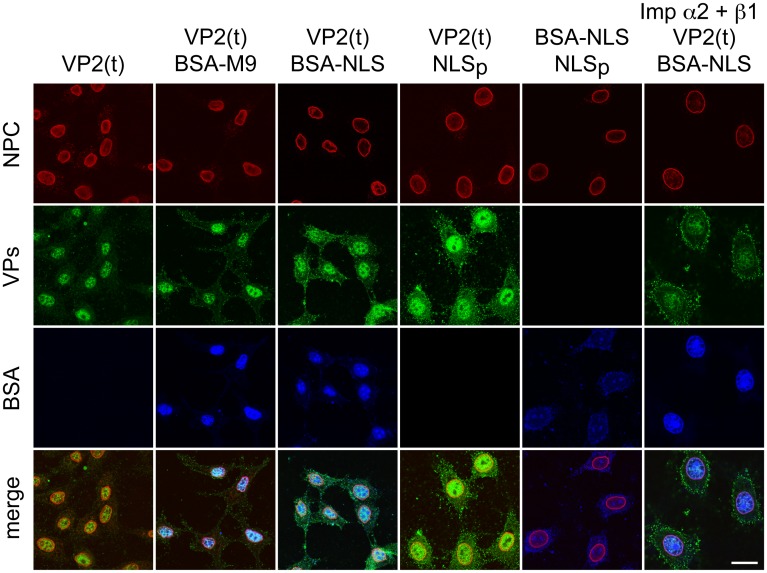
It is unknown whether the mammalian cell cycle could impact the assembly of viruses maturing in the nucleus. We addressed this question using MVM, a reference member of the icosahedral ssDNA nuclear parvoviruses, which requires cell proliferation to infect by mechanisms partly understood. Constitutively expressed MVM capsid subunits (VPs) accumulated in the cytoplasm of mouse and human fibroblasts synchronized at G0, G1, and G1/S transition. Upon arrest release, VPs translocated to the nucleus as cells entered S phase, at efficiencies relying on cell origin and arrest method, and immediately assembled into capsids. In synchronously infected cells, the consecutive virus life cycle steps (gene expression, proteins nuclear translocation, capsid assembly, genome replication and encapsidation) proceeded tightly coupled to cell cycle progression from G0/G1 through S into G2 phase. However, a DNA synthesis stress caused by thymidine irreversibly disrupted virus life cycle, as VPs became increasingly retained in the cytoplasm hours post-stress, forming empty capsids in mouse fibroblasts, thereby impairing encapsidation of the nuclear viral DNA replicative intermediates. Synchronously infected cells subjected to density-arrest signals while traversing early S phase also blocked VPs transport, resulting in a similar misplaced cytoplasmic capsid assembly in mouse fibroblasts. In contrast, thymidine and density arrest signals deregulating virus assembly neither perturbed nuclear translocation of the NS1 protein nor viral genome replication occurring under S/G2 cycle arrest. An underlying mechanism of cell cycle control was identified in the nuclear translocation of phosphorylated VPs trimeric assembly intermediates, which accessed a non-conserved route distinct from the importin α2/β1 and transportin pathways. The exquisite cell cycle-dependence of parvovirus nuclear capsid assembly conforms a novel paradigm of time and functional coupling between cellular and virus life cycles. This junction may determine the characteristic parvovirus tropism for proliferative and cancer cells, and its disturbance could critically contribute to persistence in host tissues.
哺乳动物细胞周期是否会影响在细胞核内成熟的病毒组装尚不清楚。我们使用微小病毒MVM(一种二十面体单链DNA核细小病毒的参考成员)来解决这个问题,该病毒需要细胞增殖才能感染,其机制部分已知。组成型表达的MVM衣壳亚基(VPs)在处于G0、G1以及G1/S转换期的小鼠和人成纤维细胞的细胞质中积累。解除阻滞释放后,随着细胞进入S期,VPs转移至细胞核,转移效率取决于细胞来源和阻滞方法,并立即组装成衣壳。在同步感染的细胞中,病毒连续的生命周期步骤(基因表达、蛋白质核转运、衣壳组装、基因组复制和衣壳化)与细胞周期从G0/G1经S期进入G2期的进程紧密耦合。然而,胸腺嘧啶核苷引起的DNA合成应激不可逆地破坏了病毒生命周期,因为应激数小时后VPs越来越多地滞留在细胞质中,在小鼠成纤维细胞中形成空衣壳,从而损害了核病毒DNA复制中间体的衣壳化。在穿越早期S期时受到密度阻滞信号作用的同步感染细胞也会阻断VPs的转运,导致小鼠成纤维细胞中出现类似的错误定位的细胞质衣壳组装。相比之下,解除调节病毒组装的胸腺嘧啶核苷和密度阻滞信号既不会干扰NS1蛋白的核转运,也不会干扰在S/G2期细胞周期阻滞情况下发生的病毒基因组复制。在磷酸化VPs三聚体组装中间体的核转运过程中确定了一种细胞周期控制的潜在机制,该中间体通过一条不同于输入蛋白α2/β1和运输蛋白途径的非保守途径进入细胞核。细小病毒核衣壳组装对细胞周期的精确依赖性符合细胞和病毒生命周期之间时间和功能耦合的新范式。这种连接可能决定了细小病毒对增殖细胞和癌细胞的特征性嗜性,其干扰可能对病毒在宿主组织中的持续存在起关键作用。