Pecze László, Blum Walter, Schwaller Beat
Anatomy, Department of Medicine, University of Fribourg, Route Albert-Gockel 1, CH-1700 Fribourg, Switzerland.
Anatomy, Department of Medicine, University of Fribourg, Route Albert-Gockel 1, CH-1700 Fribourg, Switzerland.
J Biol Chem. 2015 Nov 20;290(47):28214-28230. doi: 10.1074/jbc.M115.663179. Epub 2015 Sep 22.
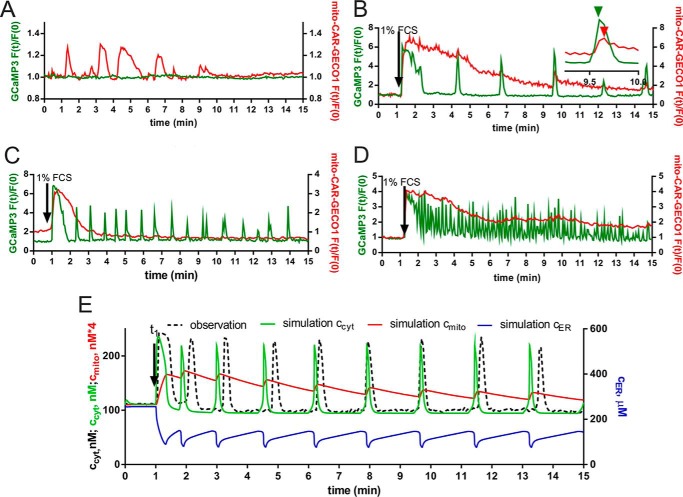
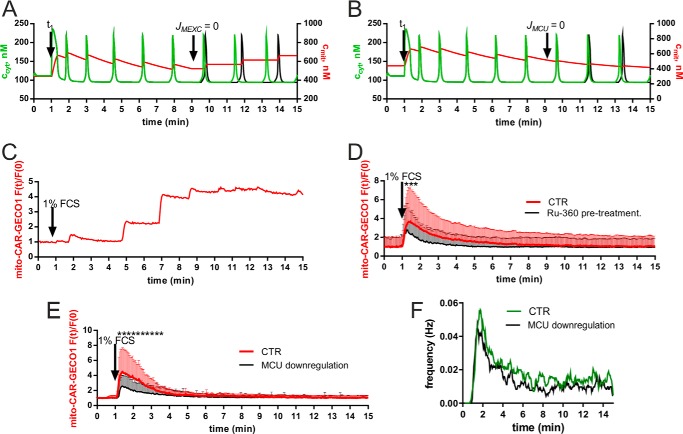
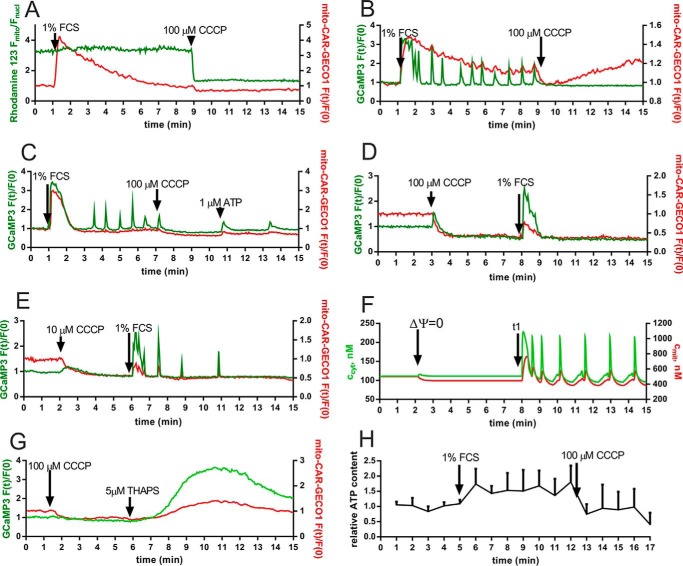
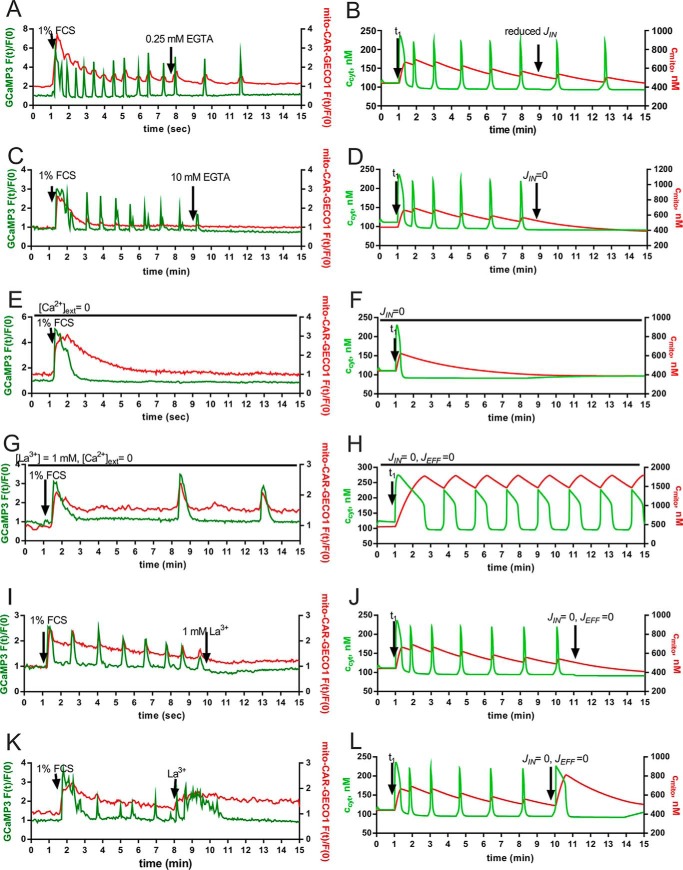
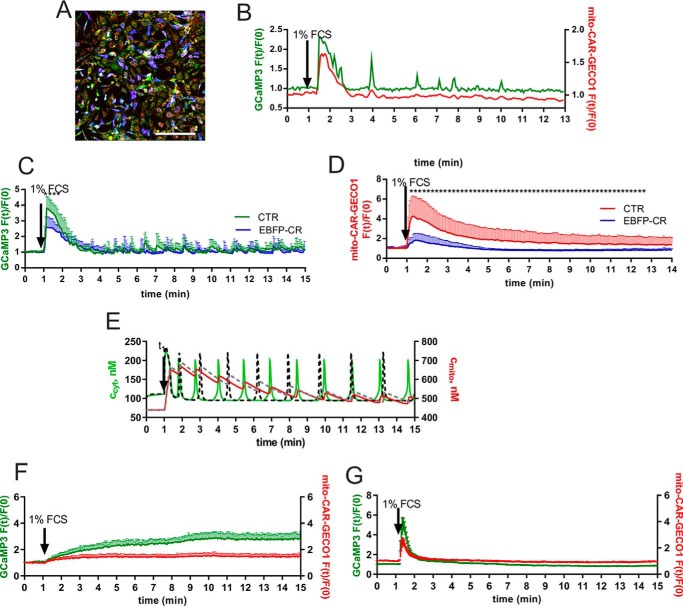
In some cell types, Ca(2+) oscillations are strictly dependent on Ca(2+) influx across the plasma membrane, whereas in others, oscillations also persist in the absence of Ca(2+) influx. We observed that, in primary mesothelial cells, the plasmalemmal Ca(2+) influx played a pivotal role. However, when the Ca(2+) transport across the plasma membrane by the "lanthanum insulation method" was blocked prior to the induction of the serum-induced Ca(2+) oscillations, mitochondrial Ca(2+) transport was found to be able to substitute for the plasmalemmal Ca(2+) exchange function, thus rendering the oscillations independent of extracellular Ca(2+). However, in a physiological situation, the Ca(2+)-buffering capacity of mitochondria was found not to be essential for Ca(2+) oscillations. Moreover, brief spontaneous Ca(2+) changes were observed in the mitochondrial Ca(2+) concentration without apparent changes in the cytosolic Ca(2+) concentration, indicating the presence of a mitochondrial autonomous Ca(2+) signaling mechanism. In the presence of calretinin, a Ca(2+)-buffering protein, the amplitude of cytosolic spikes during oscillations was decreased, and the amount of Ca(2+) ions taken up by mitochondria was reduced. Thus, the increased calretinin expression observed in mesothelioma cells and in certain colon cancer might be correlated to the increased resistance of these tumor cells to proapoptotic/pronecrotic signals. We identified and characterized (experimentally and by modeling) three Ca(2+) shuttling pathways in primary mesothelial cells during Ca(2+) oscillations: Ca(2+) shuttled between (i) the endoplasmic reticulum (ER) and mitochondria, (ii) the ER and the extracellular space, and (iii) the ER and cytoplasmic Ca(2+) buffers.
在某些细胞类型中,Ca(2+)振荡严格依赖于Ca(2+)通过质膜的内流,而在其他细胞类型中,即使没有Ca(2+)内流,振荡仍会持续。我们观察到,在原代间皮细胞中,质膜Ca(2+)内流起关键作用。然而,当通过“镧隔离法”阻断血清诱导的Ca(2+)振荡诱导前质膜上的Ca(2+)转运时,发现线粒体Ca(2+)转运能够替代质膜Ca(2+)交换功能,从而使振荡独立于细胞外Ca(2+)。然而,在生理情况下,发现线粒体的Ca(2+)缓冲能力对于Ca(2+)振荡并非必不可少。此外,在线粒体Ca(2+)浓度中观察到短暂的自发Ca(2+)变化,而胞质Ca(2+)浓度没有明显变化,这表明存在线粒体自主Ca(2+)信号机制。在存在钙结合蛋白(一种Ca(2+)缓冲蛋白)的情况下,振荡期间胞质尖峰的幅度降低,线粒体摄取的Ca(2+)离子量减少。因此,在间皮瘤细胞和某些结肠癌中观察到的钙结合蛋白表达增加可能与这些肿瘤细胞对促凋亡/促坏死信号的抗性增加有关。我们在Ca(2+)振荡期间在原代间皮细胞中鉴定并表征了(通过实验和建模)三种Ca(2+)穿梭途径:Ca(2+)在(i)内质网(ER)和线粒体之间、(ii)ER和细胞外空间之间以及(iii)ER和细胞质Ca(2+)缓冲液之间穿梭。