Gursoy-Yuzugullu Ozge, House Nealia, Price Brendan D
Department of Radiation Oncology, Dana-Farber Cancer Institute, 450 Brookline Avenue, Boston, MA 02132, USA.
Department of Radiation Oncology, Dana-Farber Cancer Institute, 450 Brookline Avenue, Boston, MA 02132, USA.
J Mol Biol. 2016 May 8;428(9 Pt B):1846-60. doi: 10.1016/j.jmb.2015.11.021. Epub 2015 Nov 26.
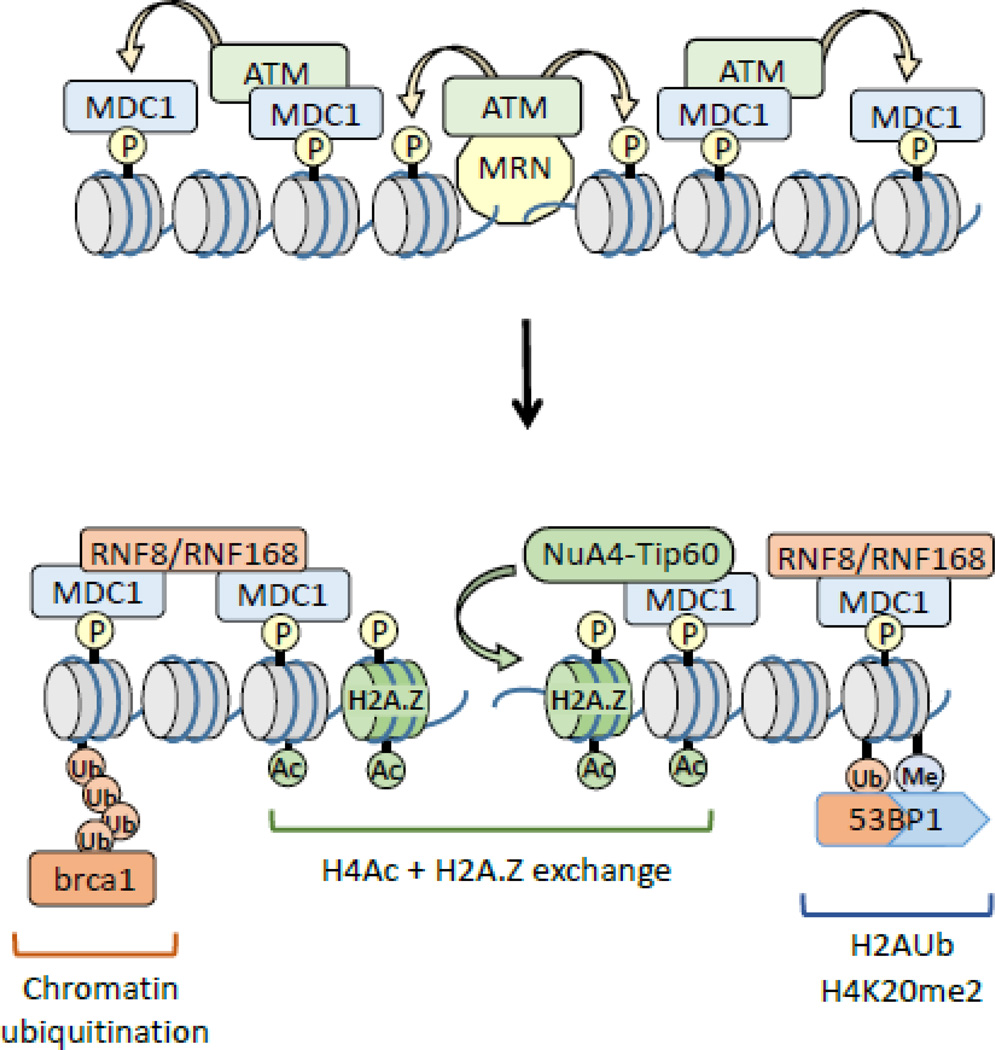
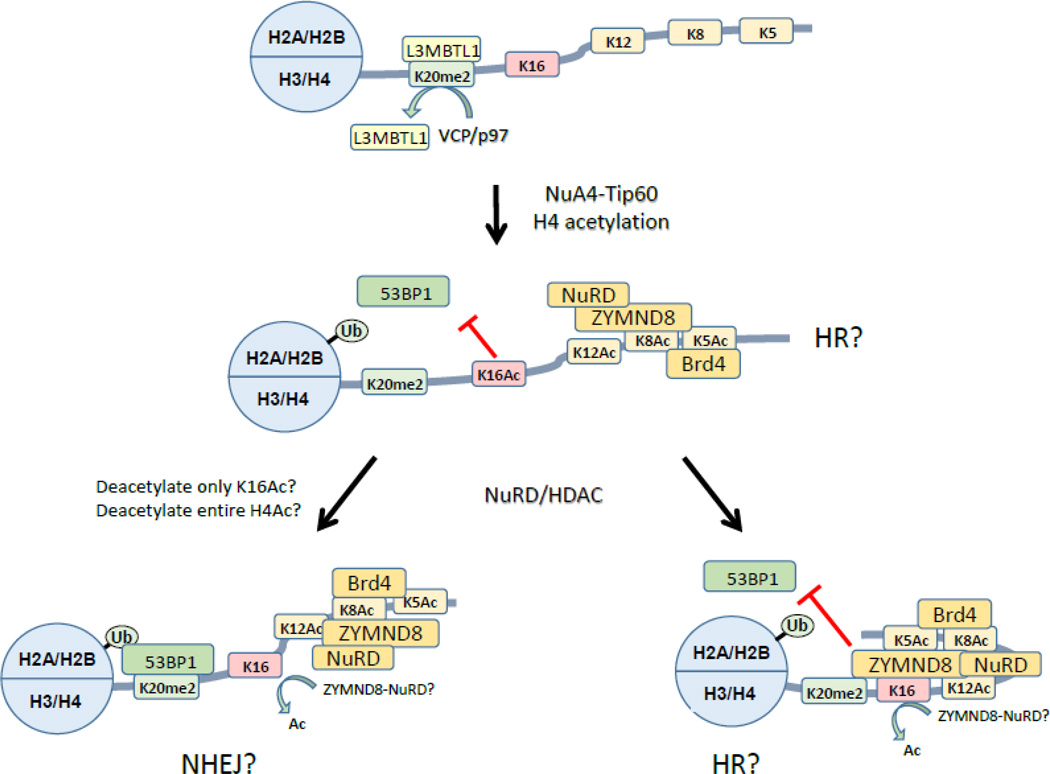
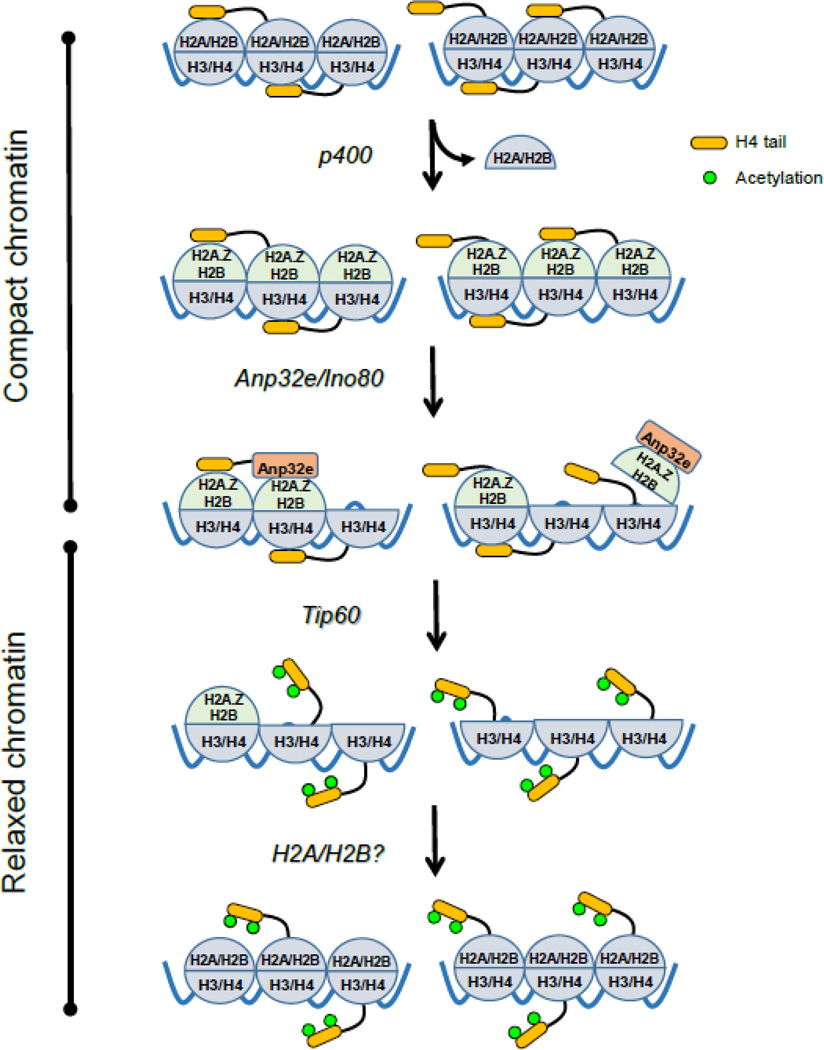
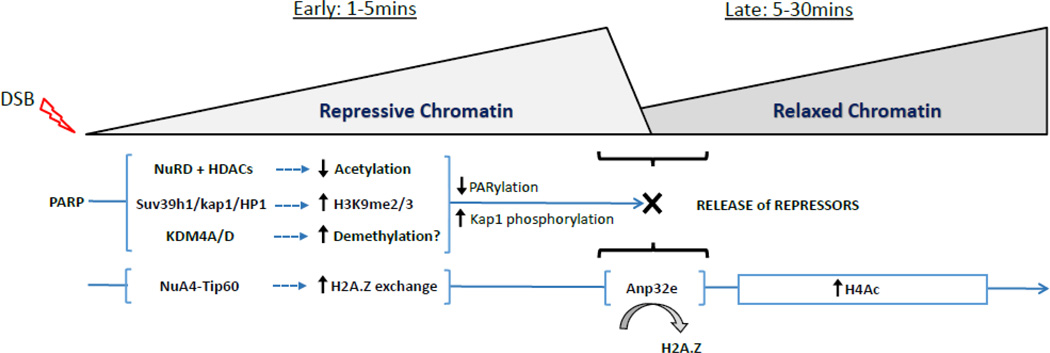
The ability of cells to detect and repair DNA double-strand breaks (DSBs) is dependent on reorganization of the surrounding chromatin structure by chromatin remodeling complexes. These complexes promote access to the site of DNA damage, facilitate processing of the damaged DNA and, importantly, are essential to repackage the repaired DNA. Here, we will review the chromatin remodeling steps that occur immediately after DSB production and that prepare the damaged chromatin template for processing by the DSB repair machinery. DSBs promote rapid accumulation of repressive complexes, including HP1, the NuRD complex, H2A.Z and histone methyltransferases at the DSB. This shift to a repressive chromatin organization may be important to inhibit local transcription and limit mobility of the break and to maintain the DNA ends in close contact. Subsequently, the repressive chromatin is rapidly dismantled through a mechanism involving dynamic exchange of the histone variant H2A.Z. H2A.Z removal at DSBs alters the acidic patch on the nucleosome surface, promoting acetylation of the H4 tail (by the NuA4-Tip60 complex) and shifting the chromatin to a more open structure. Further, H2A.Z removal promotes chromatin ubiquitination and recruitment of additional DSB repair proteins to the break. Modulation of the nucleosome surface and nucleosome function during DSB repair therefore plays a vital role in processing of DNA breaks. Further, the nucleosome surface may function as a central hub during DSB repair, directing specific patterns of histone modification, recruiting DNA repair proteins and modulating chromatin packing during processing of the damaged DNA template.
细胞检测和修复DNA双链断裂(DSB)的能力取决于染色质重塑复合物对周围染色质结构的重组。这些复合物促进对DNA损伤位点的访问,便于处理受损的DNA,并且重要的是,对于重新包装修复后的DNA至关重要。在这里,我们将回顾DSB产生后立即发生的染色质重塑步骤,这些步骤为DSB修复机制处理受损的染色质模板做好准备。DSB促进包括HP1、NuRD复合物、H2A.Z和组蛋白甲基转移酶在内的抑制性复合物在DSB处快速积累。这种向抑制性染色质组织的转变对于抑制局部转录、限制断裂的移动性以及保持DNA末端紧密接触可能很重要。随后,抑制性染色质通过一种涉及组蛋白变体H2A.Z动态交换的机制迅速解体。在DSB处去除H2A.Z会改变核小体表面的酸性斑块,促进H4尾巴的乙酰化(由NuA4-Tip60复合物介导),并使染色质转变为更开放的结构。此外,去除H2A.Z会促进染色质泛素化,并将额外的DSB修复蛋白招募到断裂处。因此,在DSB修复过程中对核小体表面和核小体功能的调节在DNA断裂的处理中起着至关重要的作用。此外,核小体表面在DSB修复过程中可能起着中心枢纽的作用,指导特定的组蛋白修饰模式,招募DNA修复蛋白,并在处理受损的DNA模板时调节染色质包装。