Klement Karolin, Luijsterburg Martijn S, Pinder Jordan B, Cena Chad S, Del Nero Victor, Wintersinger Christopher M, Dellaire Graham, van Attikum Haico, Goodarzi Aaron A
Robson DNA Science Centre, Southern Alberta Cancer Research Institute; and Department of Biochemistry and Molecular Biology and Department of Oncology, Cumming School of Medicine; University of Calgary, Calgary, Alberta T2N 4N1, Canada Robson DNA Science Centre, Southern Alberta Cancer Research Institute; and Department of Biochemistry and Molecular Biology and Department of Oncology, Cumming School of Medicine; University of Calgary, Calgary, Alberta T2N 4N1, Canada Robson DNA Science Centre, Southern Alberta Cancer Research Institute; and Department of Biochemistry and Molecular Biology and Department of Oncology, Cumming School of Medicine; University of Calgary, Calgary, Alberta T2N 4N1, Canada.
Department of Human Genetics, Leiden University Medical Centre, 2333 ZC Leiden, Netherlands.
J Cell Biol. 2014 Dec 22;207(6):717-33. doi: 10.1083/jcb.201405077.
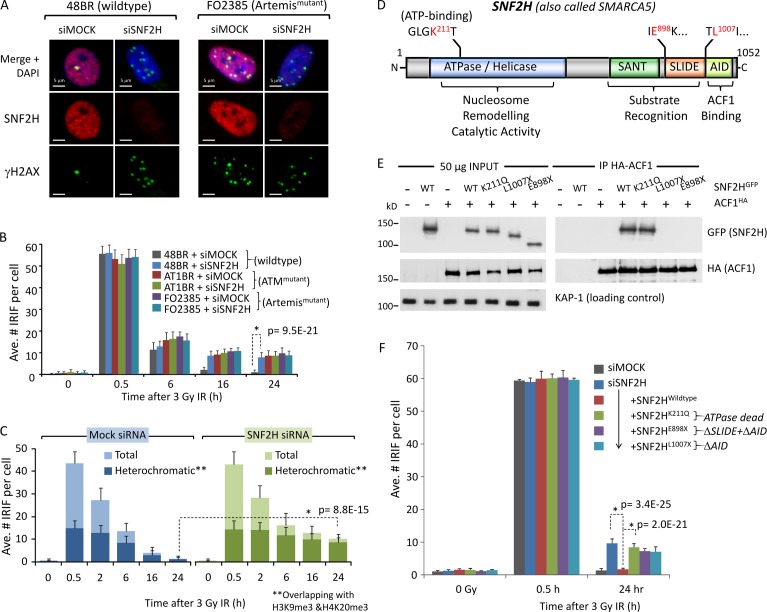
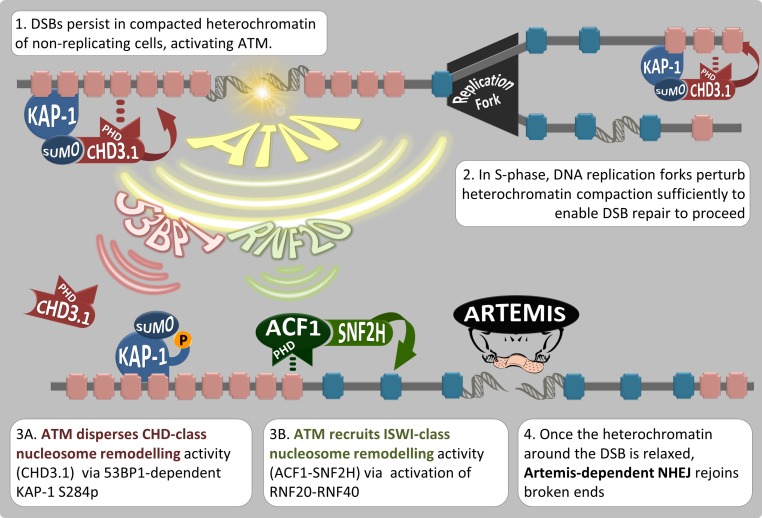
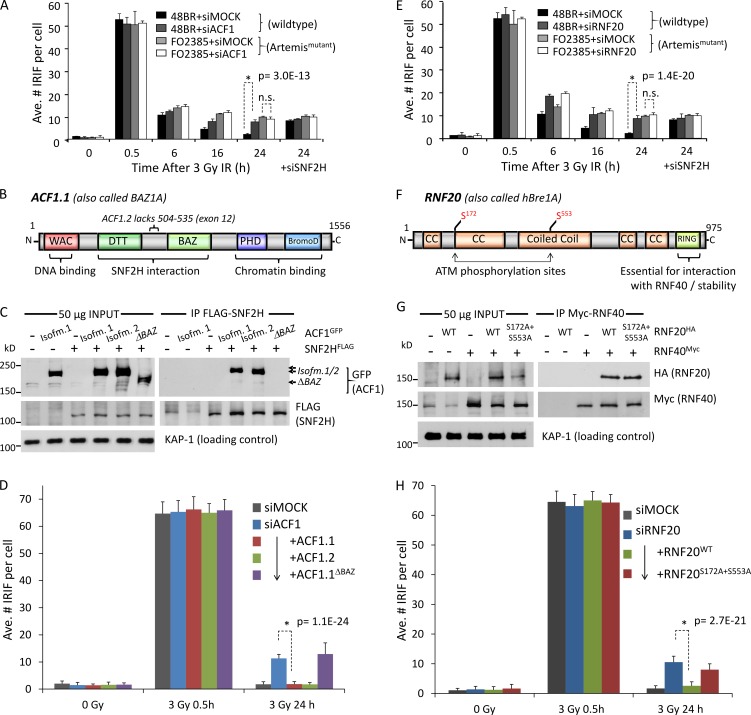
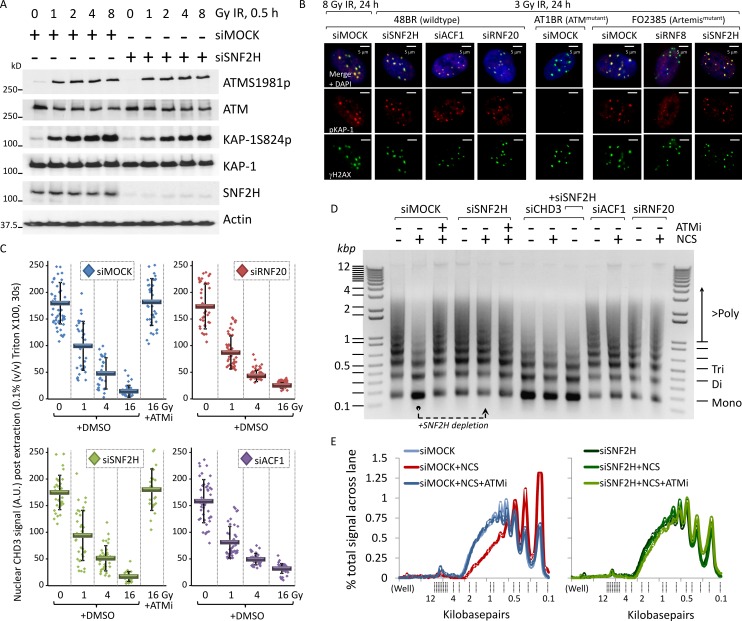
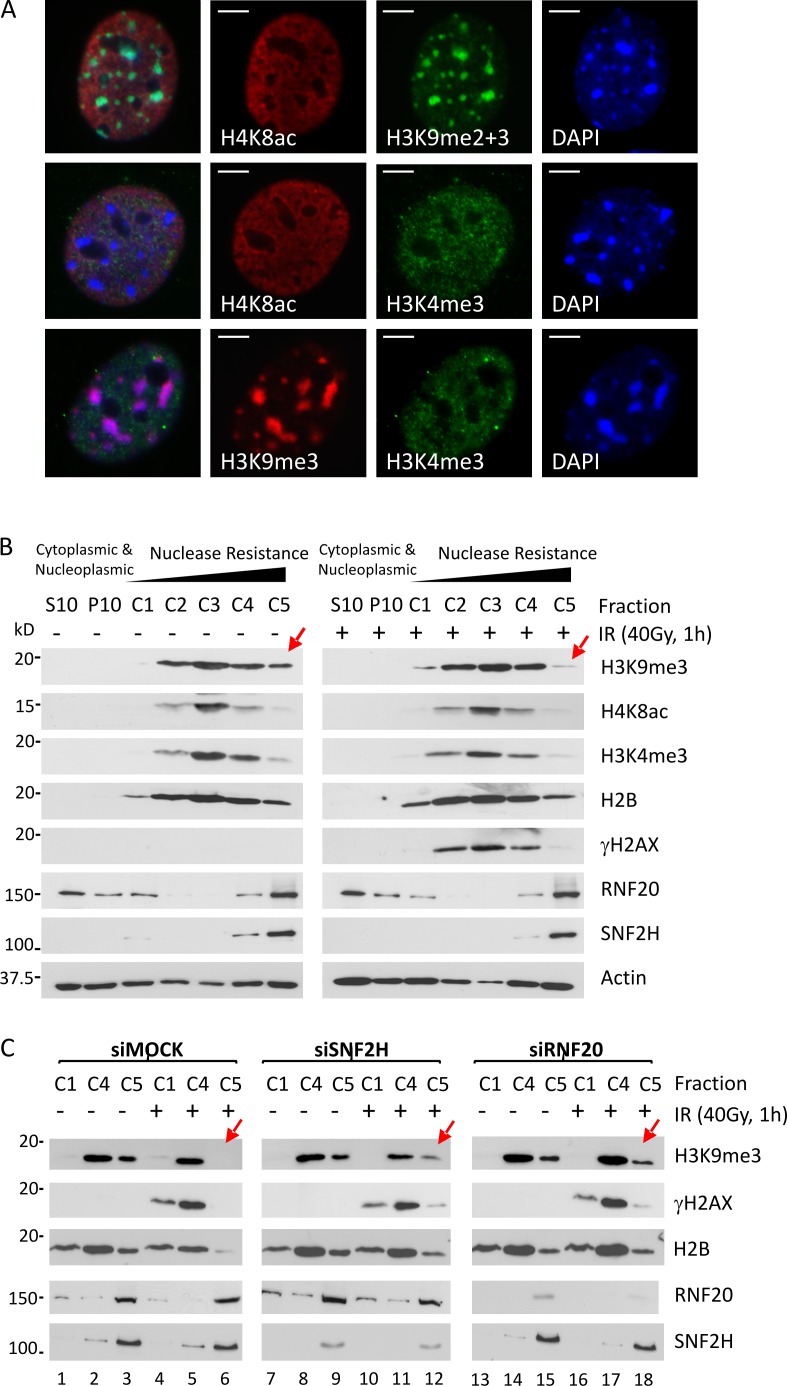
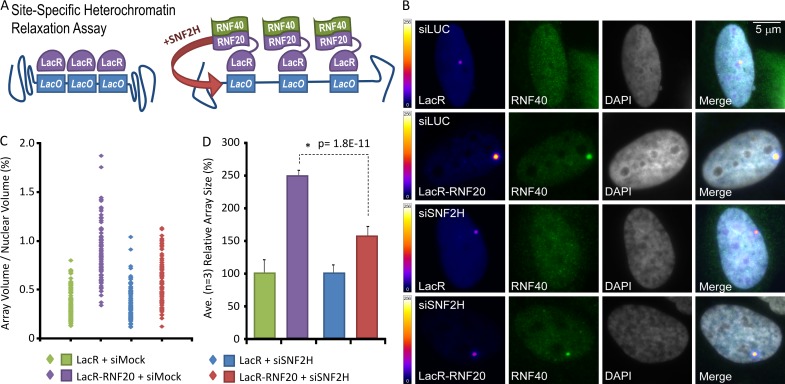
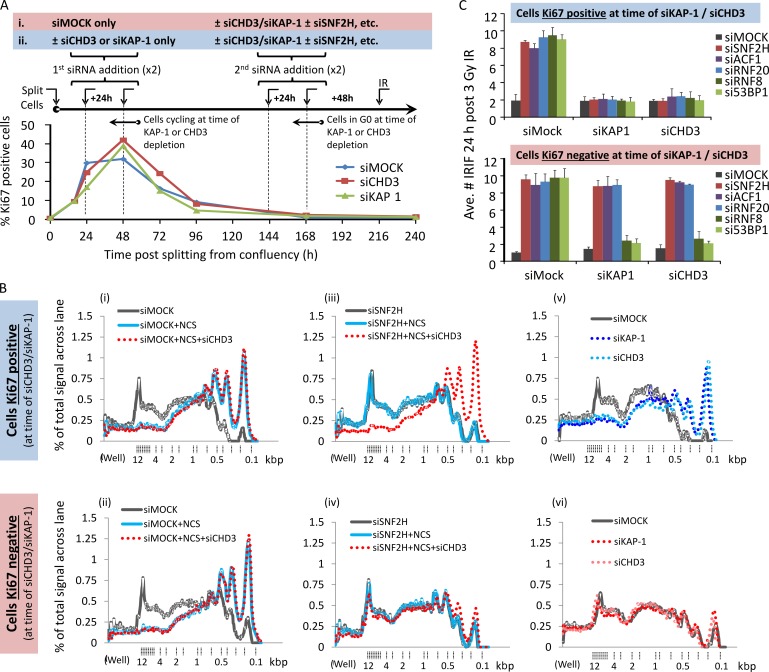
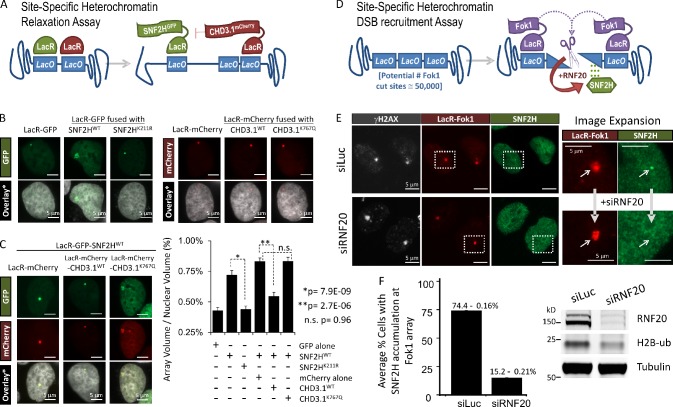
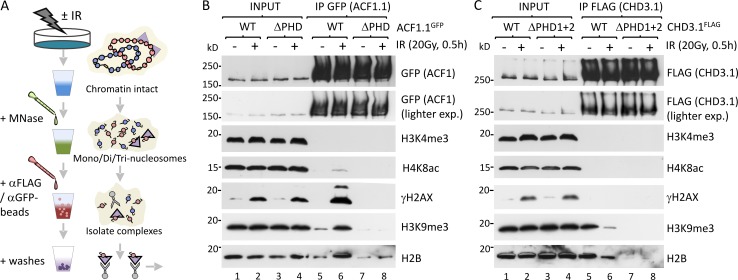
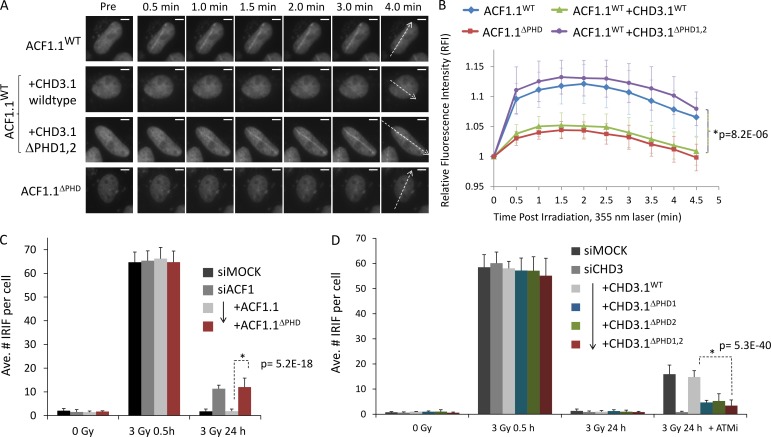
Heterochromatin is a barrier to DNA repair that correlates strongly with elevated somatic mutation in cancer. CHD class II nucleosome remodeling activity (specifically CHD3.1) retained by KAP-1 increases heterochromatin compaction and impedes DNA double-strand break (DSB) repair requiring Artemis. This obstruction is alleviated by chromatin relaxation via ATM-dependent KAP-1S824 phosphorylation (pKAP-1) and CHD3.1 dispersal from heterochromatic DSBs; however, how heterochromatin compaction is actually adjusted after CHD3.1 dispersal is unknown. In this paper, we demonstrate that Artemis-dependent DSB repair in heterochromatin requires ISWI (imitation switch)-class ACF1-SNF2H nucleosome remodeling. Compacted chromatin generated by CHD3.1 after DNA replication necessitates ACF1-SNF2H-mediated relaxation for DSB repair. ACF1-SNF2H requires RNF20 to bind heterochromatic DSBs, underlies RNF20-mediated chromatin relaxation, and functions downstream of pKAP-1-mediated CHD3.1 dispersal to enable DSB repair. CHD3.1 and ACF1-SNF2H display counteractive activities but similar histone affinities (via the plant homeodomains of CHD3.1 and ACF1), which we suggest necessitates a two-step dispersal and recruitment system regulating these opposing chromatin remodeling activities during DSB repair.
异染色质是DNA修复的障碍,与癌症中体细胞突变的增加密切相关。KAP-1保留的II类CHD核小体重塑活性(特别是CHD3.1)会增加异染色质的压缩,并阻碍需要Artemis的DNA双链断裂(DSB)修复。通过ATM依赖性KAP-1 S824磷酸化(pKAP-1)使染色质松弛以及CHD3.1从异染色质DSB处散开可缓解这种阻碍;然而,CHD3.1散开后异染色质压缩实际上是如何调节的尚不清楚。在本文中,我们证明异染色质中依赖Artemis的DSB修复需要ISWI(模仿开关)类ACF1-SNF2H核小体重塑。DNA复制后由CHD3.1产生的紧密染色质需要ACF1-SNF2H介导的松弛来进行DSB修复。ACF1-SNF2H需要RNF20结合异染色质DSB,是RNF20介导的染色质松弛的基础,并且在pKAP-1介导的CHD3.1散开的下游起作用以实现DSB修复。CHD3.1和ACF1-SNF2H表现出相反的活性,但具有相似的组蛋白亲和力(通过CHD3.1和ACF1的植物同源结构域),我们认为这需要一个两步的散开和招募系统来在DSB修复过程中调节这些相反的染色质重塑活性。