Yu Xianxian, Duan Xiaoshan, Zhang Rui, Fu Xuehao, Ye Lingling, Kong Hongzhi, Xu Guixia, Shan Hongyan
State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of SciencesBeijing, China; University of Chinese Academy of SciencesBeijing, China.
State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences Beijing, China.
Front Plant Sci. 2016 May 2;7:598. doi: 10.3389/fpls.2016.00598. eCollection 2016.
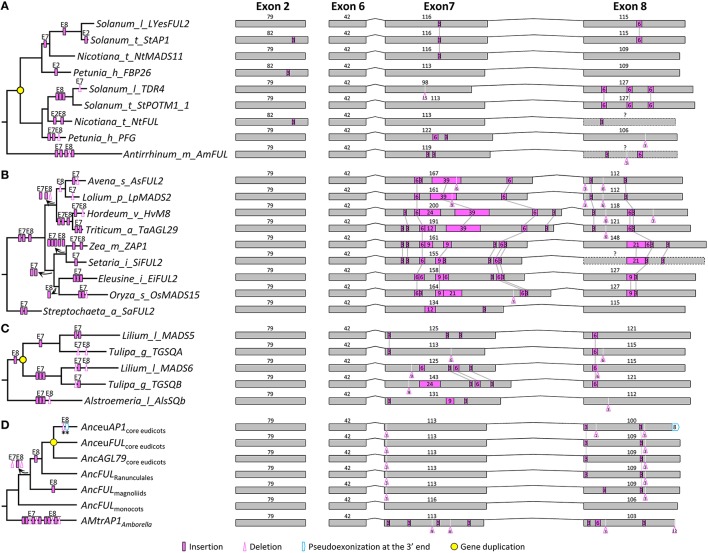
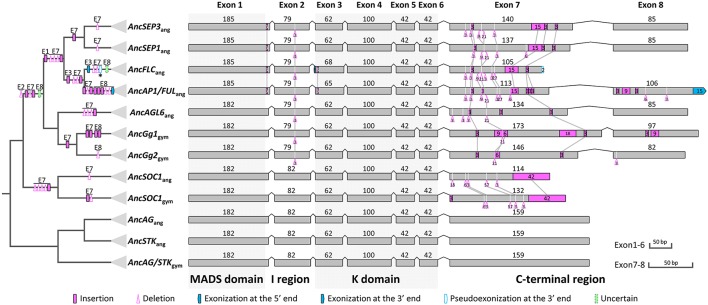
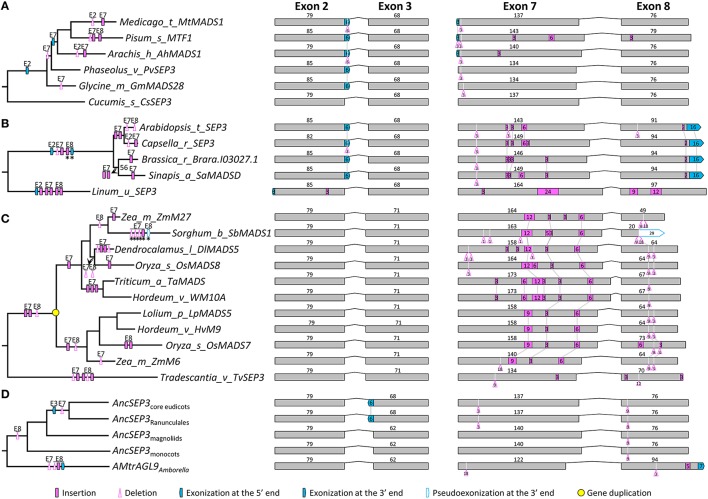
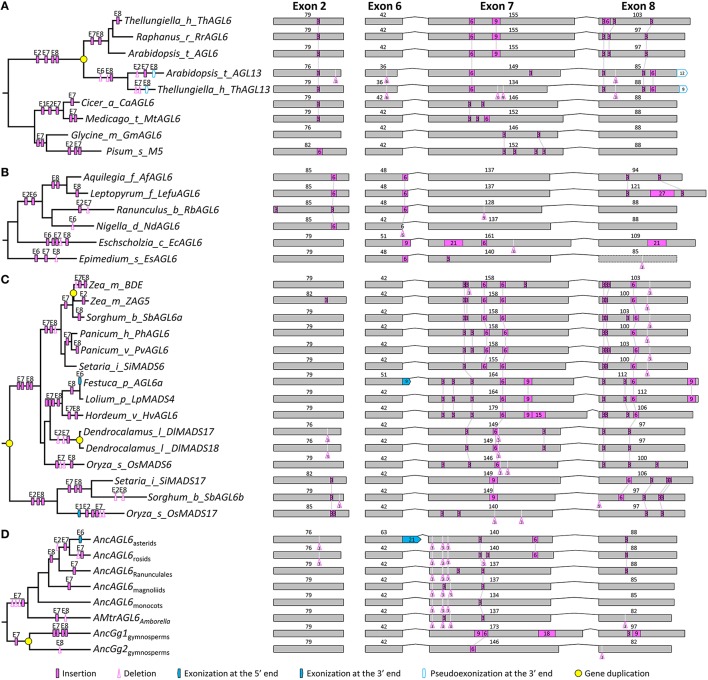
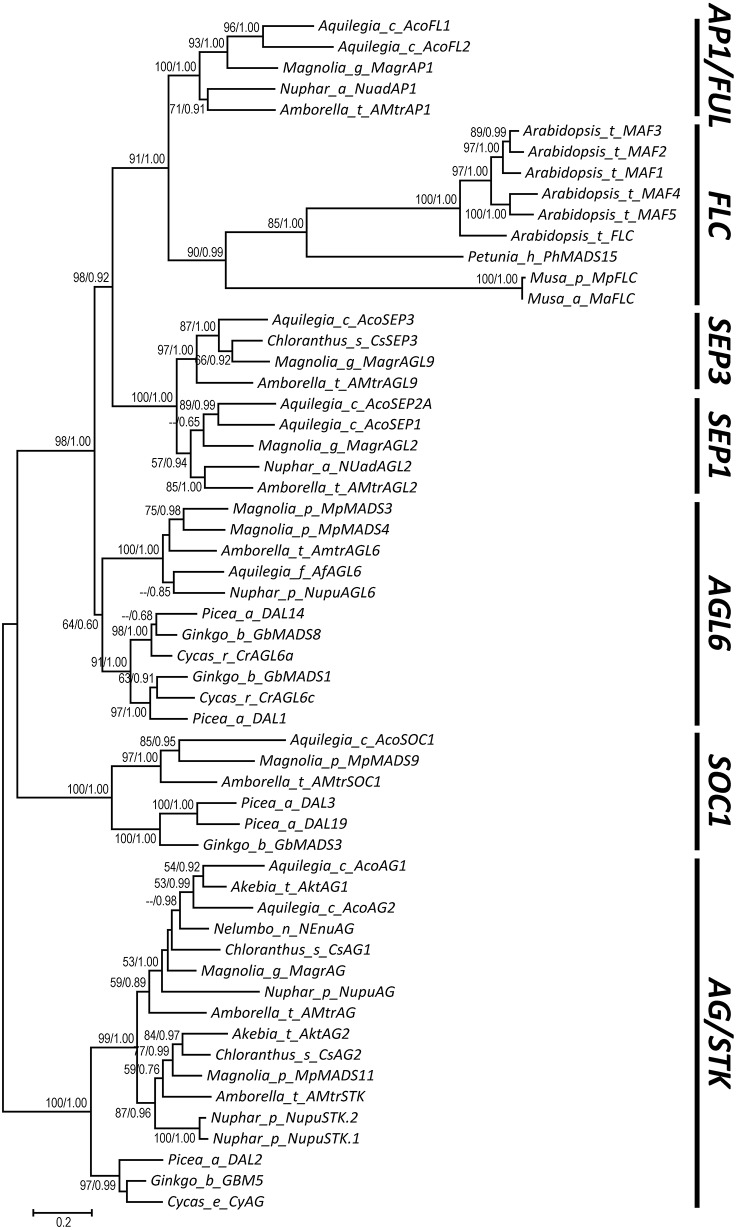
AP1/FUL, SEP, AGL6, and FLC subfamily genes play important roles in flower development. The phylogenetic relationships among them, however, have been controversial, which impedes our understanding of the origin and functional divergence of these genes. One possible reason for the controversy may be the problems caused by changes in the exon-intron structure of genes, which, according to recent studies, may generate non-homologous sites and hamper the homology-based sequence alignment. In this study, we first performed exon-by-exon alignments of these and three outgroup subfamilies (SOC1, AG, and STK). Phylogenetic trees reconstructed based on these matrices show improved resolution and better congruence with species phylogeny. In the context of these phylogenies, we traced evolutionary changes of exon-intron structures in each subfamily. We found that structural changes have occurred frequently following gene duplication and speciation events. Notably, exons 7 and 8 (if present) suffered more structural changes than others. With the knowledge of exon-intron structural changes, we generated more reasonable alignments containing all the focal subfamilies. The resulting trees showed that the SEP subfamily is sister to the monophyletic group formed by AP1/FUL and FLC subfamily genes and that the AGL6 subfamily forms a sister group to the three abovementioned subfamilies. Based on this topology, we inferred the evolutionary history of exon-intron structural changes among different subfamilies. Particularly, we found that the eighth exon originated before the divergence of AP1/FUL, FLC, SEP, and AGL6 subfamilies and degenerated in the ancestral FLC-like gene. These results provide new insights into the origin and evolution of the AP1/FUL, FLC, SEP, and AGL6 subfamilies.
AP1/FUL、SEP、AGL6和FLC亚家族基因在花发育过程中发挥着重要作用。然而,它们之间的系统发育关系一直存在争议,这阻碍了我们对这些基因的起源和功能分化的理解。争议的一个可能原因可能是基因外显子-内含子结构变化所导致的问题,根据最近的研究,这种变化可能会产生非同源位点并妨碍基于同源性的序列比对。在本研究中,我们首先对这些亚家族以及三个外类群亚家族(SOC1、AG和STK)进行了逐个外显子的比对。基于这些矩阵重建的系统发育树显示出更高的分辨率,并且与物种系统发育具有更好的一致性。在这些系统发育的背景下,我们追踪了每个亚家族中外显子-内含子结构的进化变化。我们发现,结构变化在基因复制和物种形成事件之后频繁发生。值得注意的是,外显子7和8(如果存在)比其他外显子经历了更多的结构变化。基于对外显子-内含子结构变化的了解,我们生成了包含所有重点亚家族的更合理的比对。由此得到的系统发育树表明,SEP亚家族是由AP1/FUL和FLC亚家族基因形成的单系群的姐妹群,而AGL6亚家族则是上述三个亚家族的姐妹群。基于这种拓扑结构,我们推断了不同亚家族之间外显子-内含子结构变化的进化历史。特别是,我们发现第八个外显子在AP1/FUL、FLC、SEP和AGL6亚家族分化之前就已出现,并在祖先的FLC样基因中退化。这些结果为AP1/FUL、FLC、SEP和AGL6亚家族的起源和进化提供了新的见解。