Kong Lingchun, Tan Li, Lv Ruitu, Shi Zhennan, Xiong Lijun, Wu Feizhen, Rabidou Kimberlie, Smith Michael, He Celestine, Zhang Lei, Qian Yanyan, Ma Duan, Lan Fei, Shi Yang, Shi Yujiang Geno
Laboratory of Epigenetics, Institutes of Biomedical Sciences, Shanghai Medical College of Fudan University, Shanghai 200032, China.
Laboratory of Epigenetics, Institutes of Biomedical Sciences, Shanghai Medical College of Fudan University, Shanghai 200032, China
Nucleic Acids Res. 2016 Oct 14;44(18):8682-8692. doi: 10.1093/nar/gkw529. Epub 2016 Jun 10.
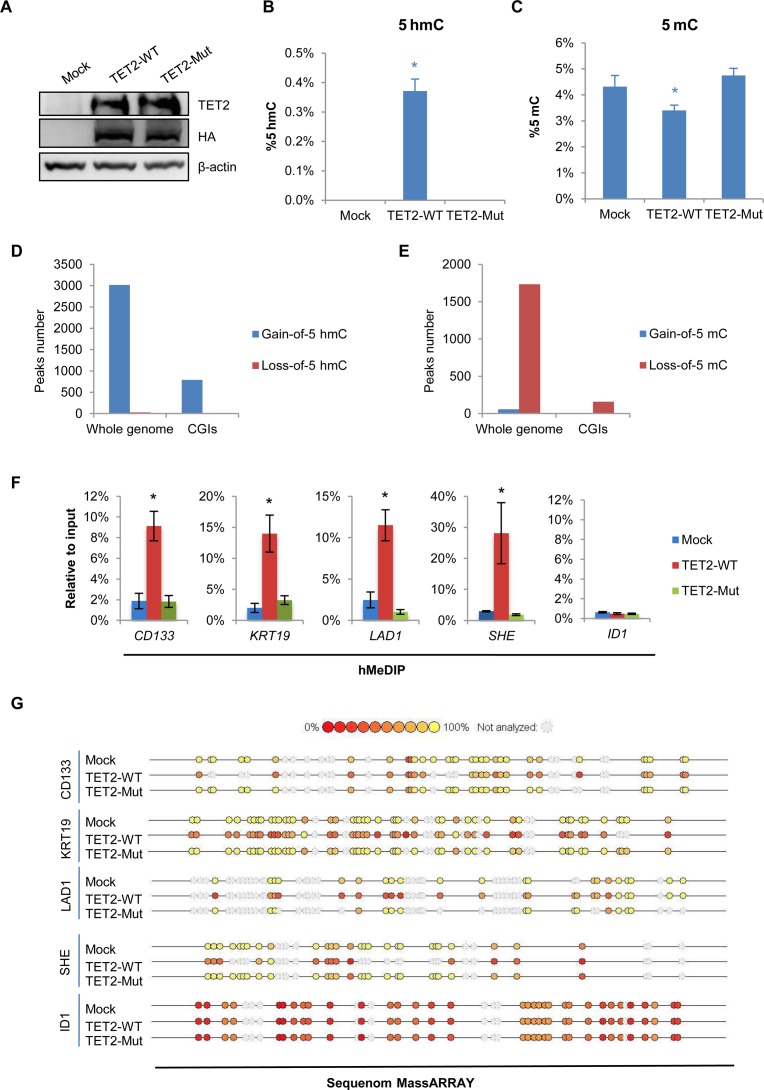
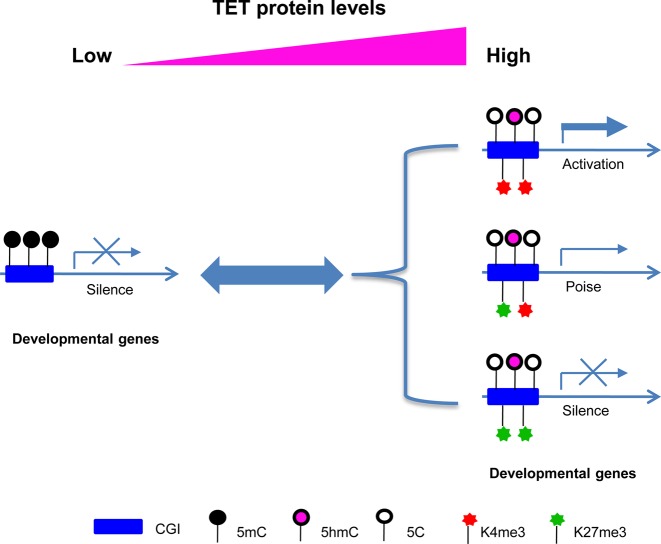
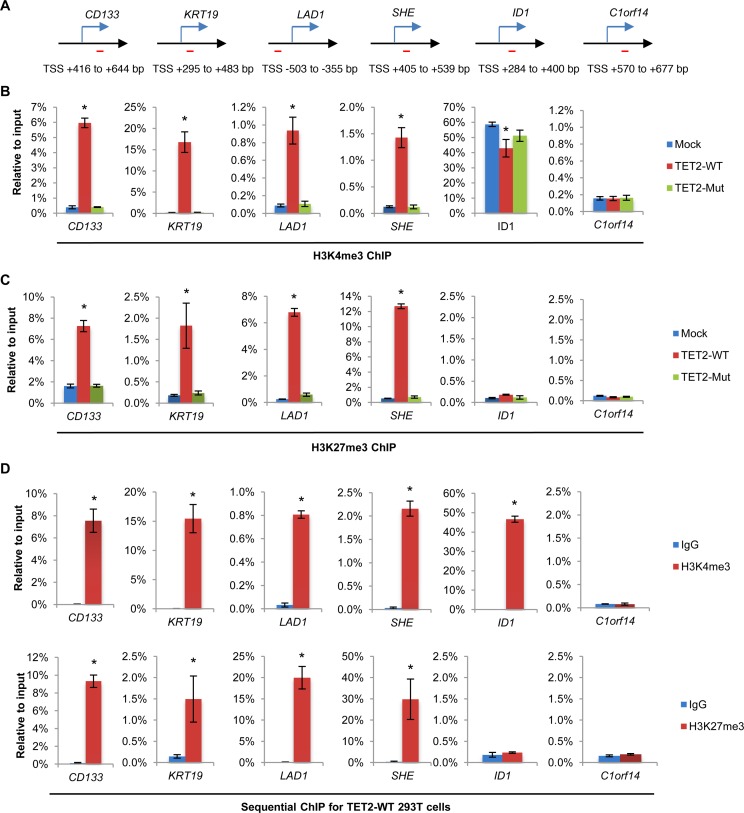
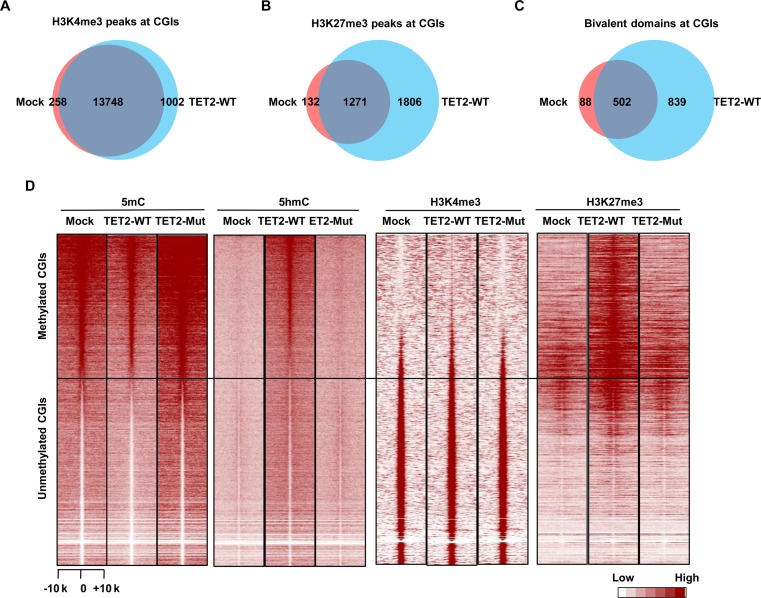
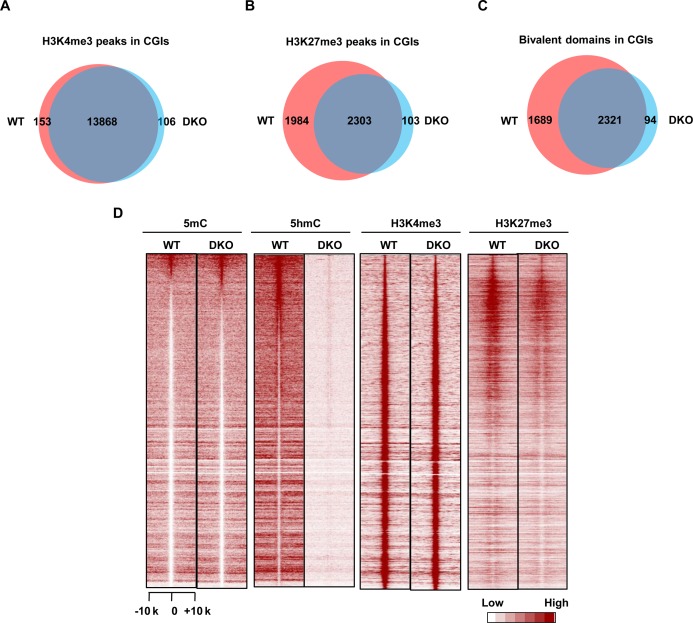
Ten Eleven Translocation (TET) protein-catalyzed 5mC oxidation not only creates novel DNA modifications, such as 5hmC, but also initiates active or passive DNA demethylation. TETs' role in the crosstalk with specific histone modifications, however, is largely elusive. Here, we show that TET2-mediated DNA demethylation plays a primary role in the de novo establishment and maintenance of H3K4me3/H3K27me3 bivalent domains underlying methylated DNA CpG islands (CGIs). Overexpression of wild type (WT), but not catalytic inactive mutant (Mut), TET2 in low-TET-expressing cells results in an increase in the level of 5hmC with accompanying DNA demethylation at a subset of CGIs. Most importantly, this alteration is sufficient in making de novo bivalent domains at these loci. Genome-wide analysis reveals that these de novo synthesized bivalent domains are largely associated with a subset of essential developmental gene promoters, which are located within CGIs and are previously silenced due to DNA methylation. On the other hand, deletion of Tet1 and Tet2 in mouse embryonic stem (ES) cells results in an apparent loss of H3K27me3 at bivalent domains, which are associated with a particular set of key developmental gene promoters. Collectively, this study demonstrates the critical role of TET proteins in regulating the crosstalk between two key epigenetic mechanisms, DNA methylation and histone methylation (H3K4me3 and H3K27me3), particularly at CGIs associated with developmental genes.
10-11易位(TET)蛋白催化的5-甲基胞嘧啶(5mC)氧化不仅能产生新的DNA修饰,如5-羟甲基胞嘧啶(5hmC),还能启动主动或被动的DNA去甲基化。然而,TET蛋白在与特定组蛋白修饰的相互作用中的作用在很大程度上仍不清楚。在这里,我们表明,TET2介导的DNA去甲基化在甲基化DNA的CpG岛(CGI)所对应的H3K4me3/H3K27me3双价结构域的从头建立和维持中起主要作用。在低TET表达细胞中过表达野生型(WT)而非催化失活突变体(Mut)的TET2,会导致5hmC水平升高,同时在一部分CGI处伴随DNA去甲基化。最重要的是,这种改变足以在这些位点从头合成双价结构域。全基因组分析表明,这些从头合成的双价结构域在很大程度上与一组必需的发育基因启动子相关,这些启动子位于CGI内,并且由于DNA甲基化先前处于沉默状态。另一方面,在小鼠胚胎干细胞(ES)中缺失Tet1和Tet2会导致双价结构域处H3K27me3明显缺失,这些双价结构域与一组特定的关键发育基因启动子相关。总的来说,这项研究证明了TET蛋白在调节两种关键表观遗传机制——DNA甲基化和组蛋白甲基化(H3K4me3和H3K27me3)之间的相互作用中起着关键作用,特别是在与发育基因相关的CGI处。