Su Hui Teo Catherine, Serwa Remigiusz A, O'Hare Peter
Section of Virology, Faculty of Medicine, Imperial College London, St Mary's Medical School, Norfolk Place, London, United Kingdom.
Department of Chemistry, Imperial College London, London, United Kingdom.
PLoS Pathog. 2016 Oct 5;12(10):e1005927. doi: 10.1371/journal.ppat.1005927. eCollection 2016 Oct.
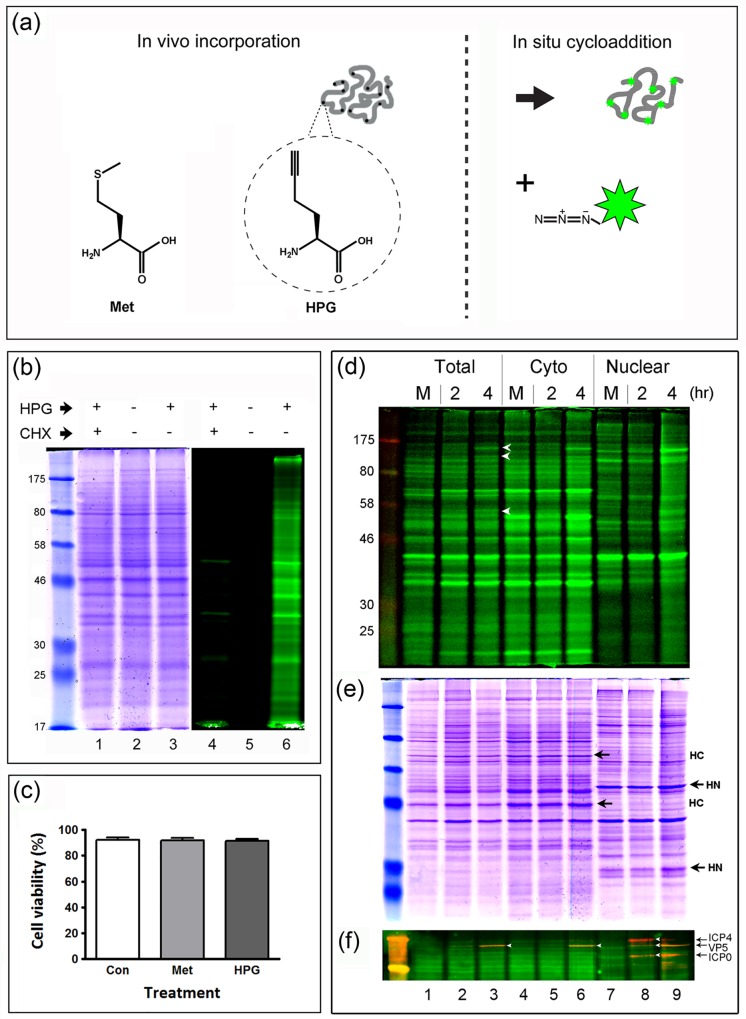
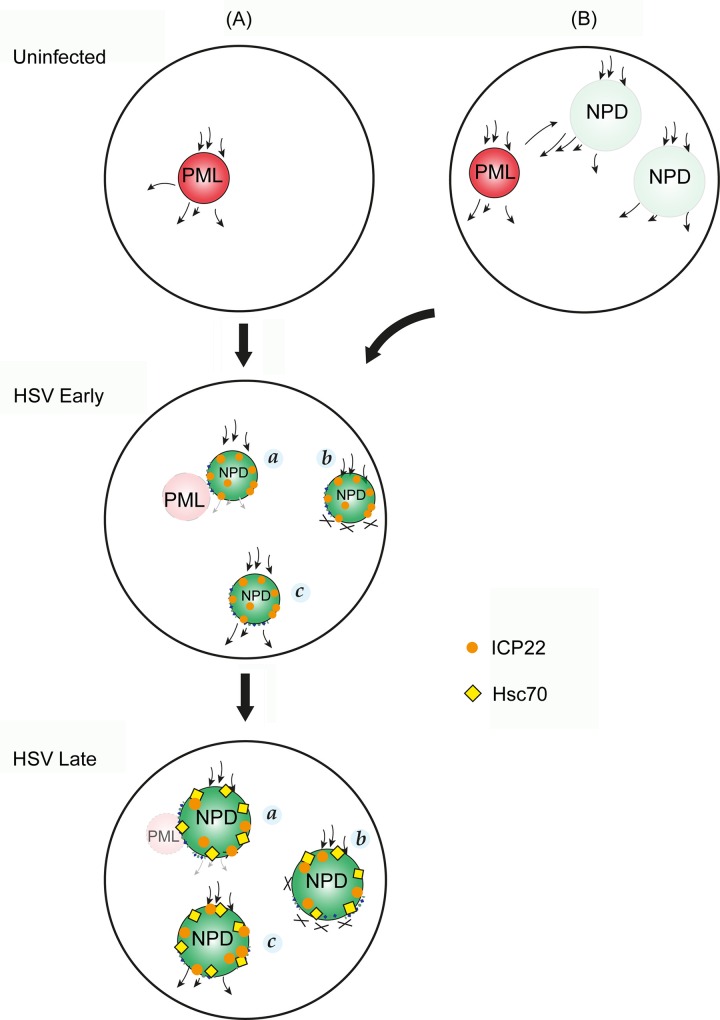
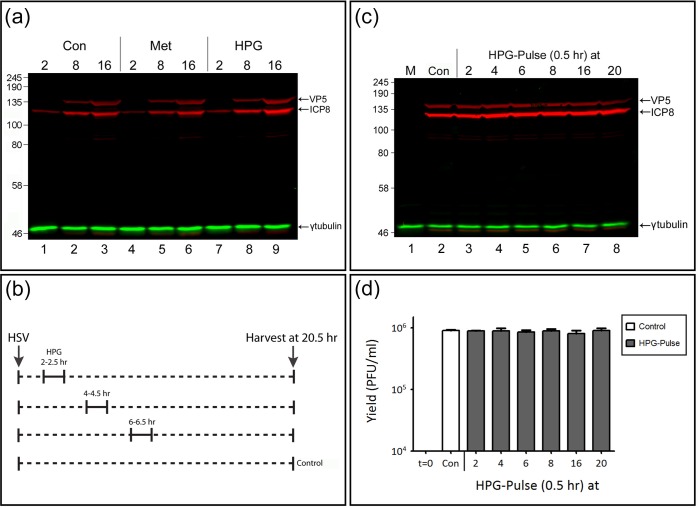
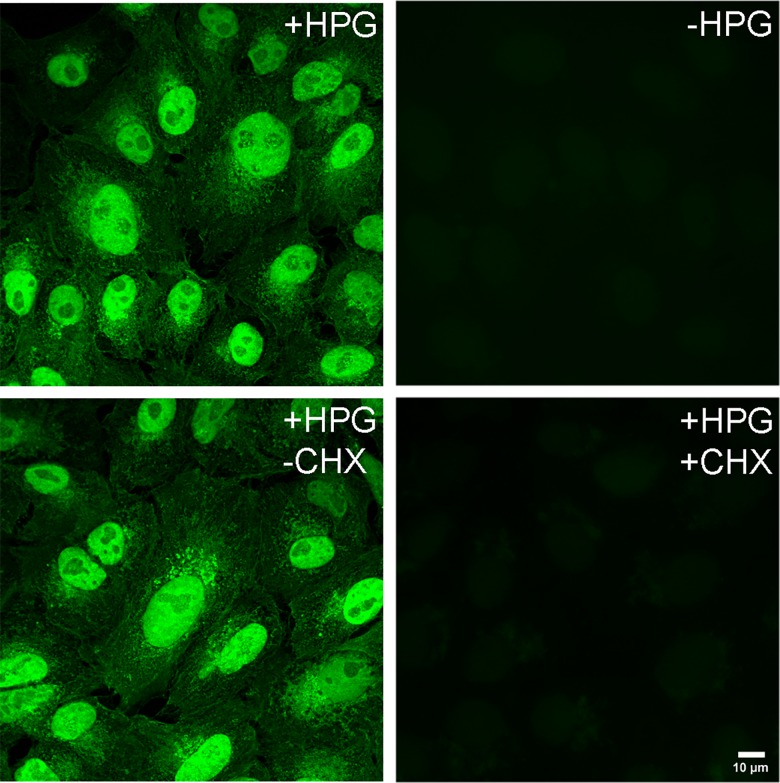
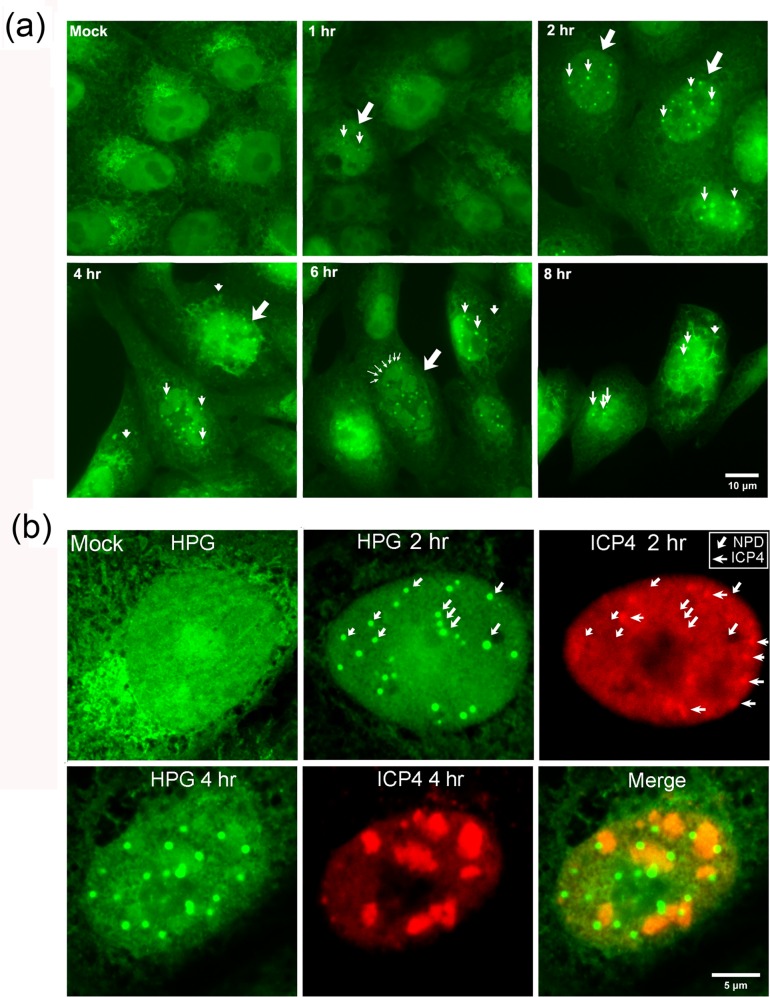
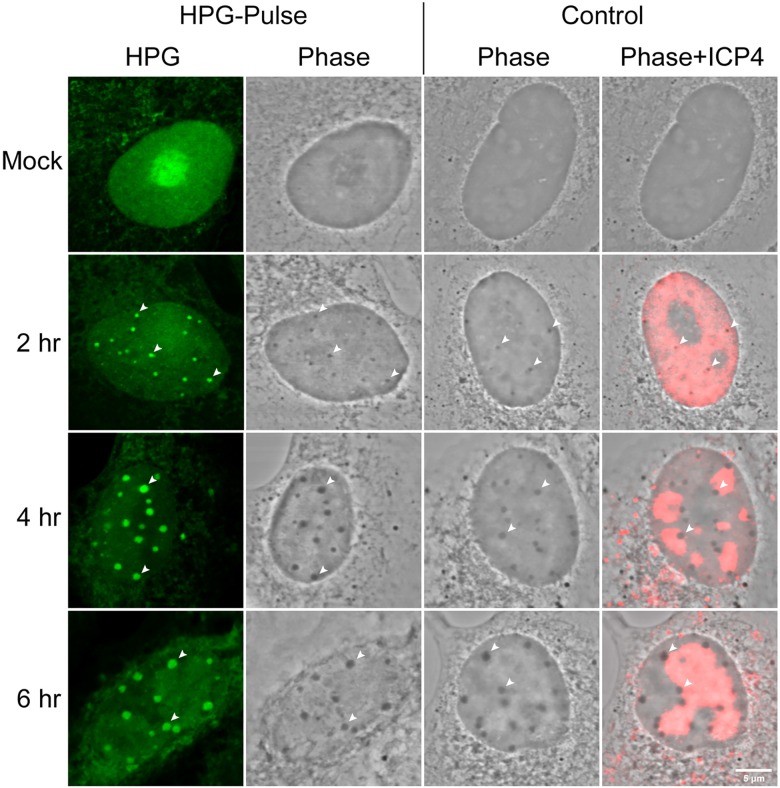
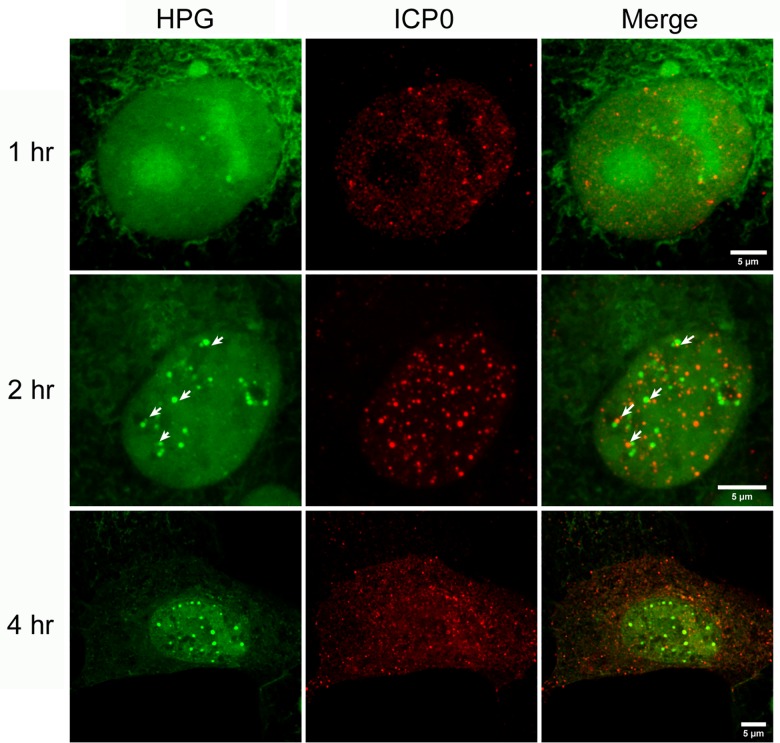
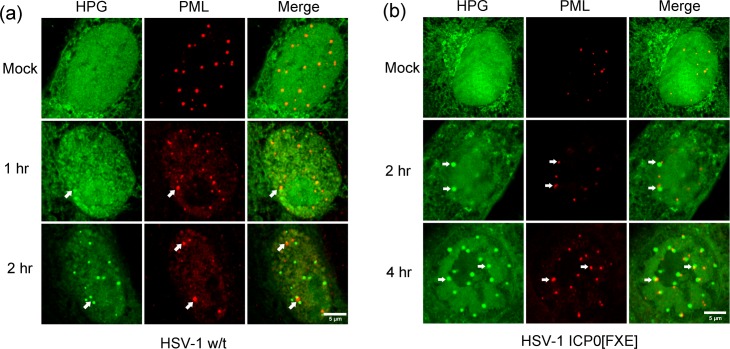
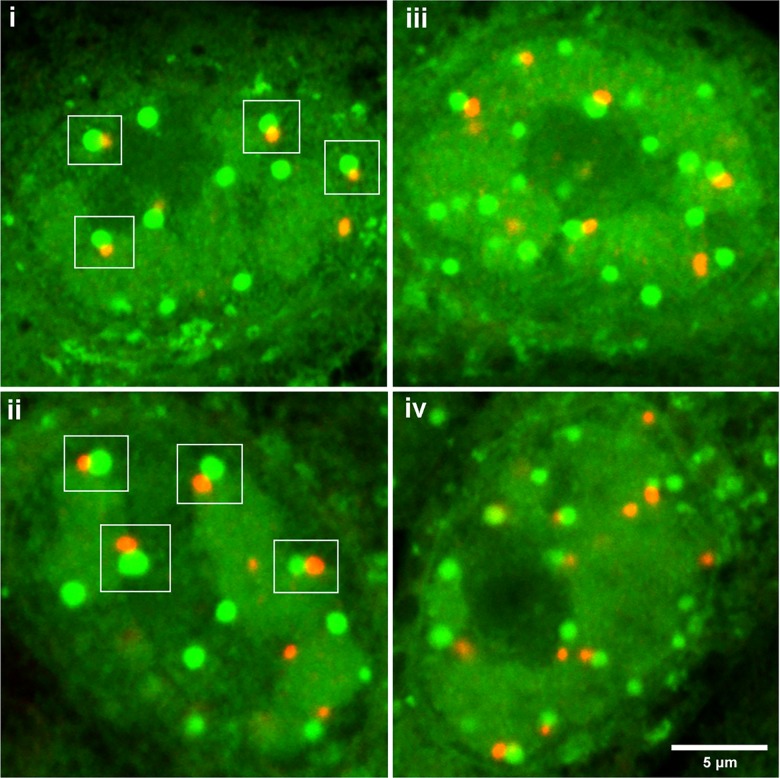
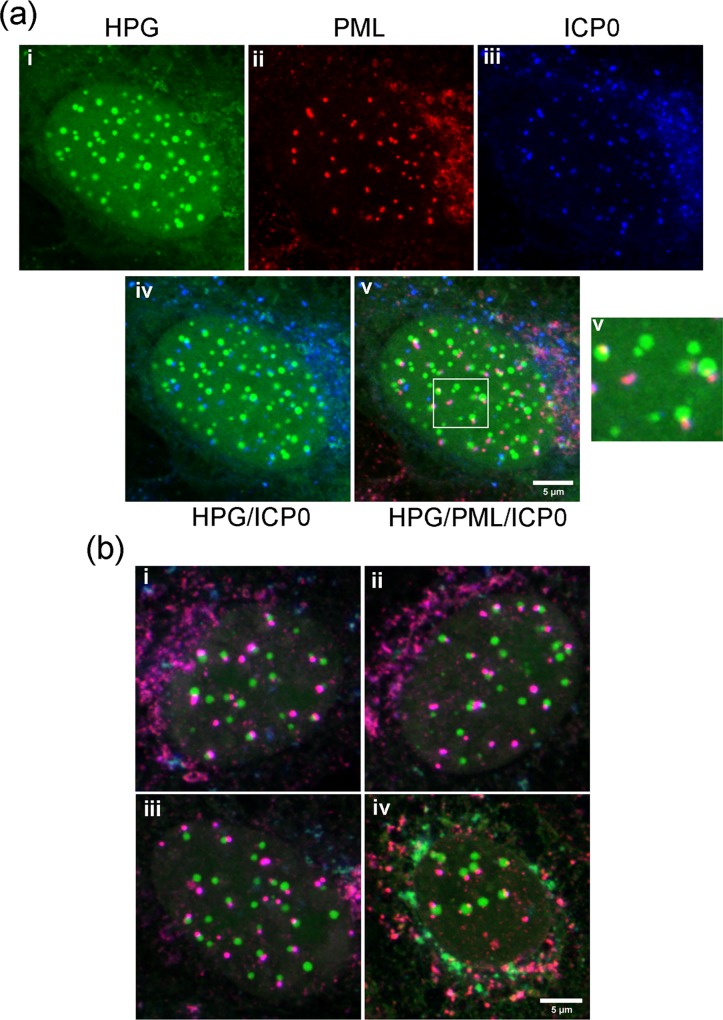
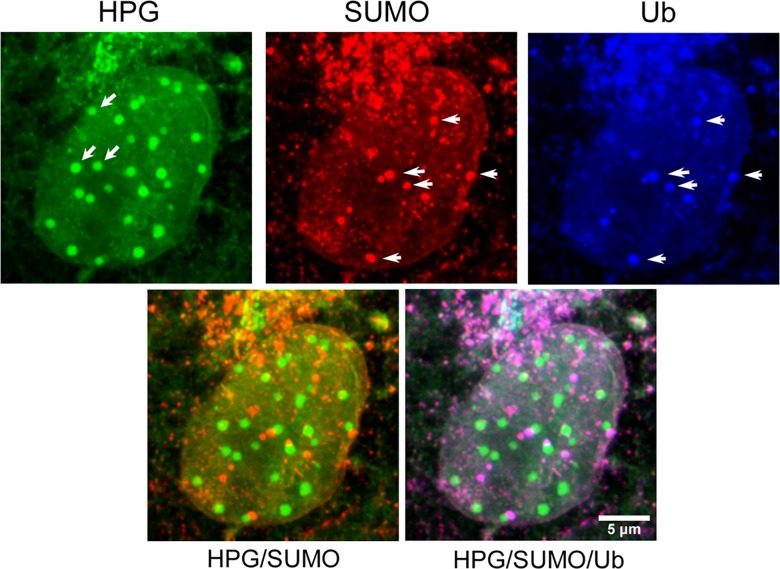
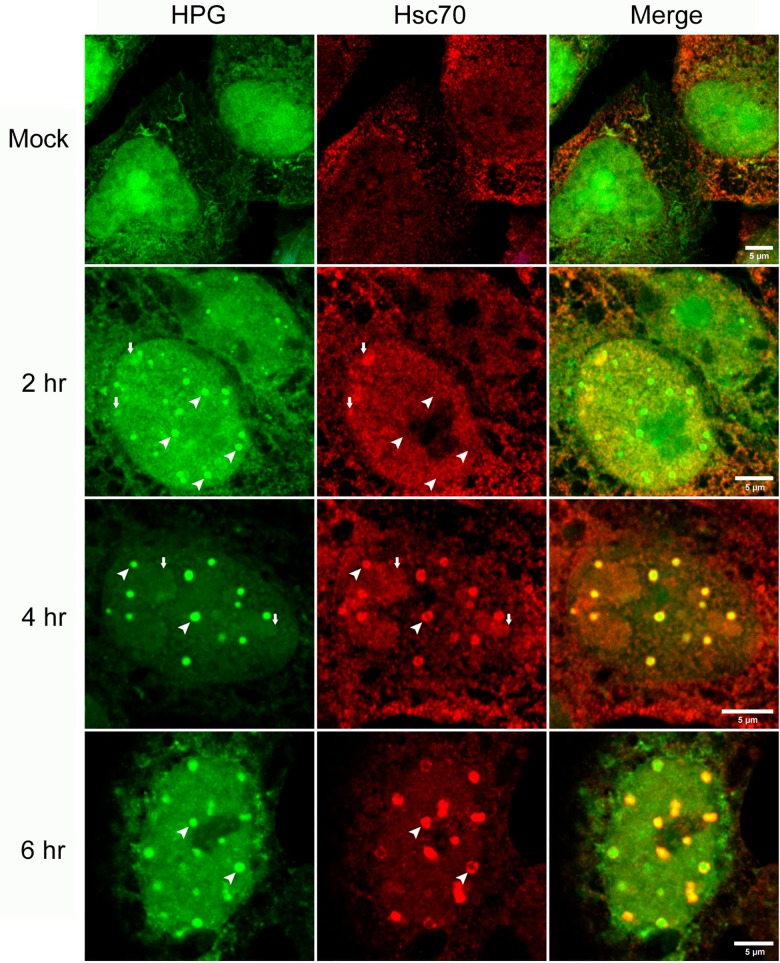
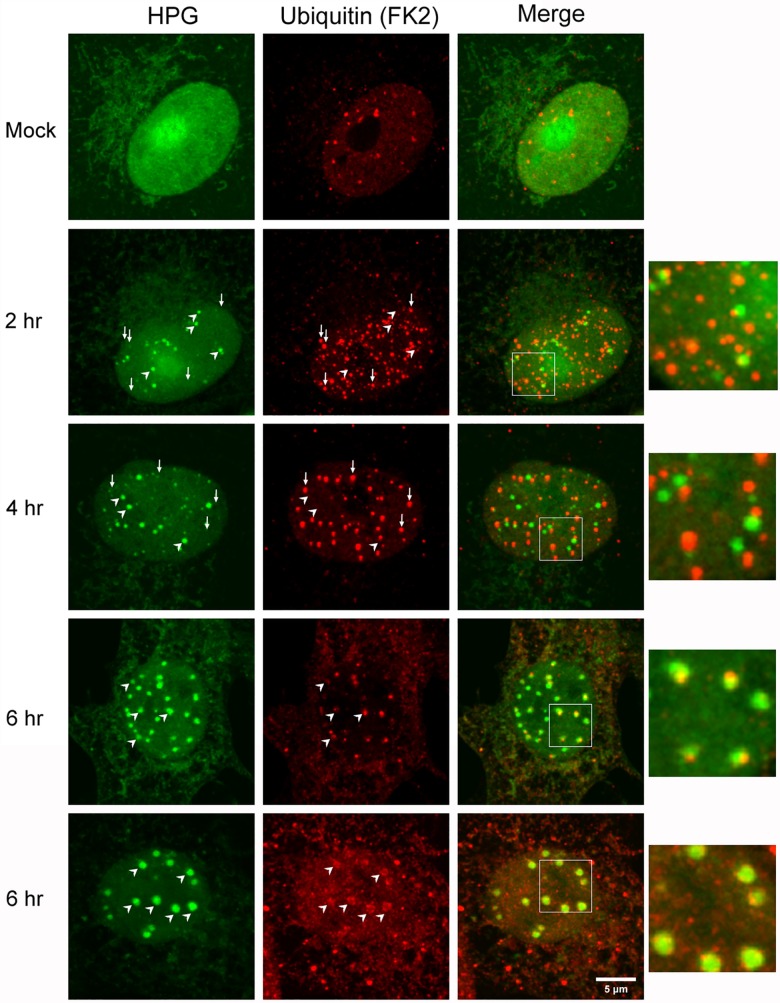
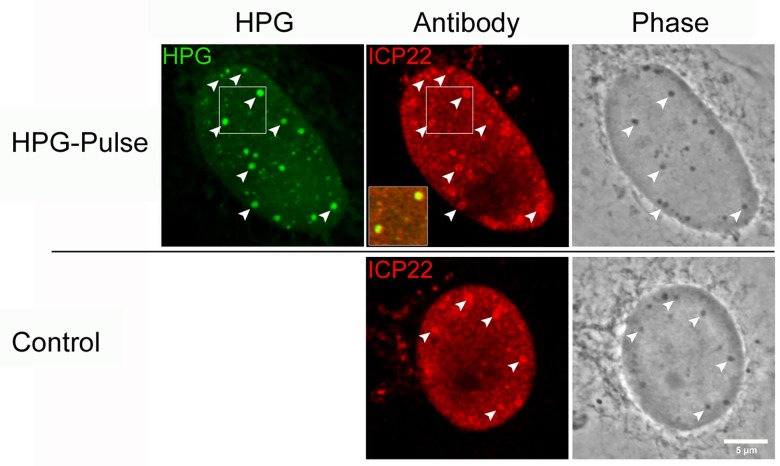
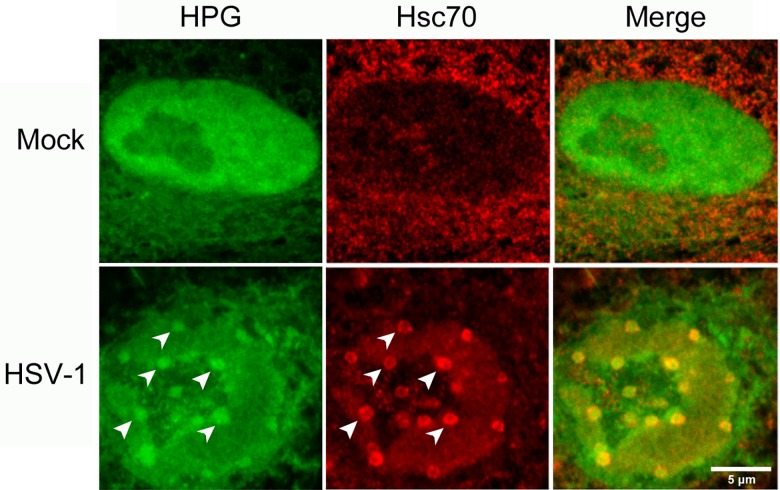
We used pulse-labeling with the methionine analogue homopropargylglycine (HPG) to investigate spatiotemporal aspects of protein synthesis during herpes simplex virus (HSV) infection. In vivo incorporation of HPG enables subsequent selective coupling of fluorochrome-capture reagents to newly synthesised proteins. We demonstrate that HPG labeling had no effect on cell viability, on accumulation of test early or late viral proteins, or on overall virus yields. HPG pulse-labeling followed by SDS-PAGE analysis confirmed incorporation into newly synthesised proteins, while parallel processing by in situ cycloaddition revealed new insight into spatiotemporal aspects of protein localisation during infection. A striking feature was the rapid accumulation of newly synthesised proteins not only in a general nuclear pattern but additionally in newly forming sub-compartments represented by small discrete foci. These newly synthesised protein domains (NPDs) were similar in size and morphology to PML domains but were more numerous, and whereas PML domains were progressively disrupted, NPDs were progressively induced and persisted. Immediate-early proteins ICP4 and ICP0 were excluded from NPDs, but using an ICP0 mutant defective in PML disruption, we show a clear spatial relationship between NPDs and PML domains with NPDs frequently forming immediately adjacent and co-joining persisting PML domains. Further analysis of location of the chaperone Hsc70 demonstrated that while NPDs formed early in infection without overt Hsc70 recruitment, later in infection Hsc70 showed pronounced recruitment frequently in a coat-like fashion around NPDs. Moreover, while ICP4 and ICP0 were excluded from NPDs, ICP22 showed selective recruitment. Our data indicate that NPDs represent early recruitment of host and viral de novo translated protein to distinct structural entities which are precursors to the previously described VICE domains involved in protein quality control in the nucleus, and reveal new features from which we propose spatially linked platforms of newly synthesised protein processing after nuclear import.
我们使用甲硫氨酸类似物高炔丙基甘氨酸(HPG)进行脉冲标记,以研究单纯疱疹病毒(HSV)感染期间蛋白质合成的时空特征。HPG在体内的掺入使得随后能够将荧光染料捕获试剂选择性地与新合成的蛋白质偶联。我们证明,HPG标记对细胞活力、测试早期或晚期病毒蛋白的积累或总体病毒产量均无影响。HPG脉冲标记后进行SDS-PAGE分析证实其掺入了新合成的蛋白质中,而通过原位环加成的平行处理揭示了感染期间蛋白质定位时空特征的新见解。一个显著特征是新合成的蛋白质不仅以一般的核模式快速积累,而且还在由小的离散病灶代表的新形成的亚区室中积累。这些新合成的蛋白质结构域(NPDs)在大小和形态上与PML结构域相似,但数量更多,并且虽然PML结构域逐渐被破坏,但NPDs逐渐被诱导并持续存在。早期即刻蛋白ICP4和ICP0被排除在NPDs之外,但使用在PML破坏方面有缺陷的ICP0突变体,我们显示NPDs与PML结构域之间存在明确的空间关系,NPDs经常紧邻并连接持续存在的PML结构域形成。伴侣蛋白Hsc70定位的进一步分析表明,虽然NPDs在感染早期形成时没有明显的Hsc70募集,但在感染后期,Hsc70显示出明显的募集,经常以类似外壳的方式围绕NPDs。此外,虽然ICP4和ICP0被排除在NPDs之外,但ICP22显示出选择性募集。我们的数据表明,NPDs代表宿主和病毒从头翻译的蛋白质早期募集到不同的结构实体,这些实体是先前描述的参与细胞核蛋白质质量控制的VICE结构域的前体,并揭示了新特征,据此我们提出核输入后新合成蛋白质加工的空间连接平台。