Department of Neuroscience and Pharmacology, Faculty of Health and Medical Sciences, University of Copenhagen, Copenhagen, Denmark.
Elife. 2016 Oct 26;5:e18805. doi: 10.7554/eLife.18805.
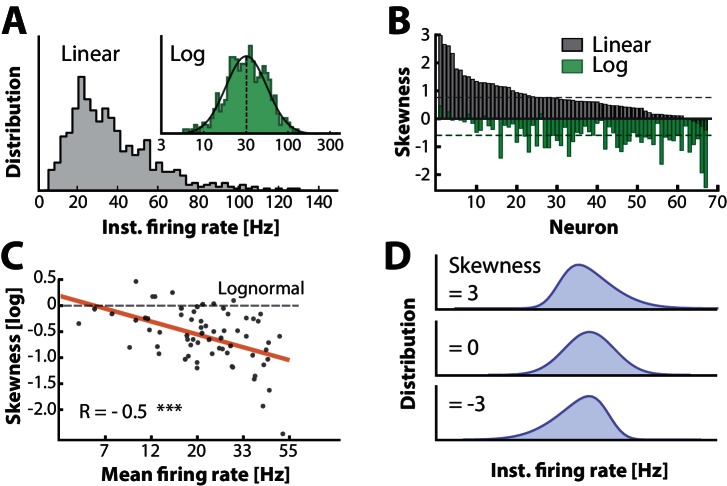
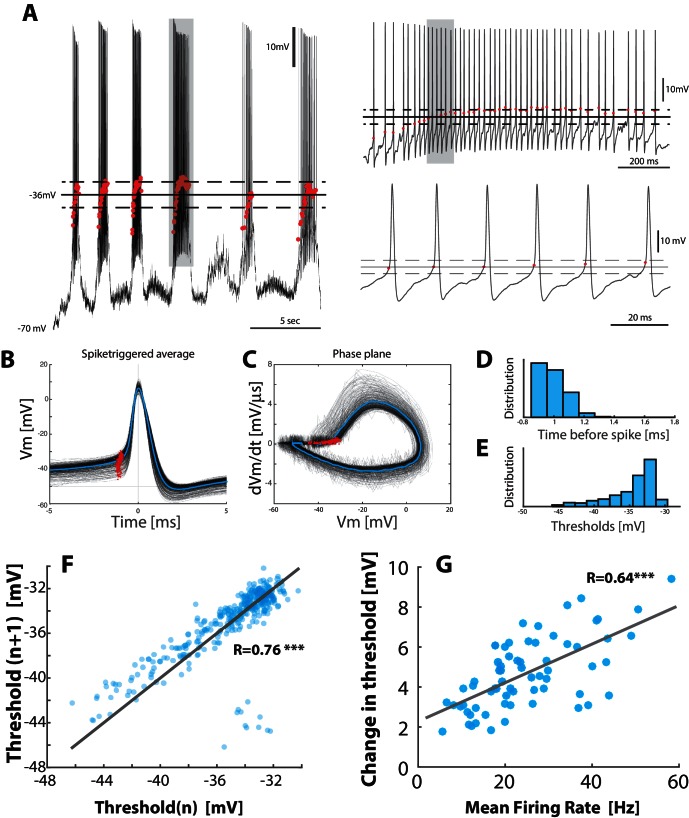
When spinal circuits generate rhythmic movements it is important that the neuronal activity remains within stable bounds to avoid saturation and to preserve responsiveness. Here, we simultaneously record from hundreds of neurons in lumbar spinal circuits of turtles and establish the neuronal fraction that operates within either a 'mean-driven' or a 'fluctuation-driven' regime. Fluctuation-driven neurons have a 'supralinear' input-output curve, which enhances sensitivity, whereas the mean-driven regime reduces sensitivity. We find a rich diversity of firing rates across the neuronal population as reflected in a lognormal distribution and demonstrate that half of the neurons spend at least 50 % of the time in the 'fluctuation-driven' regime regardless of behavior. Because of the disparity in input-output properties for these two regimes, this fraction may reflect a fine trade-off between stability and sensitivity in order to maintain flexibility across behaviors.
当脊髓回路产生有节奏的运动时,重要的是神经元活动保持在稳定的范围内,以避免饱和并保持响应能力。在这里,我们同时记录了海龟腰髓回路中的数百个神经元,并确定了在“均值驱动”或“波动驱动”模式下工作的神经元部分。波动驱动的神经元具有“超线性”的输入-输出曲线,从而提高了灵敏度,而均值驱动的模式则降低了灵敏度。我们发现神经元群体中的放电率具有丰富的多样性,这反映在对数正态分布中,并证明无论行为如何,有一半的神经元至少有 50%的时间处于“波动驱动”模式。由于这两个模式的输入-输出特性存在差异,这个比例可能反映了在稳定性和灵敏度之间的精细权衡,以在各种行为中保持灵活性。