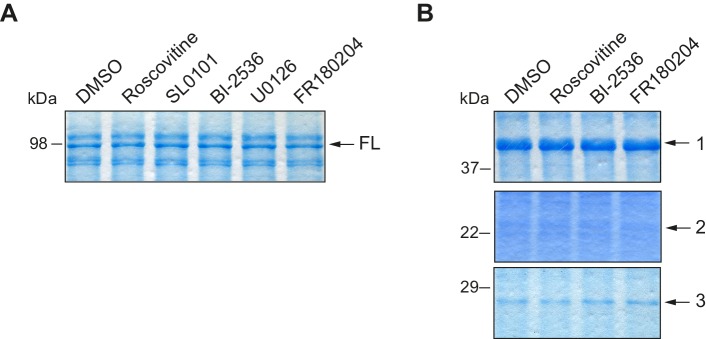
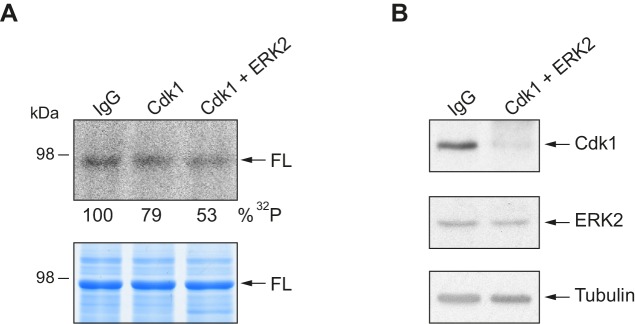
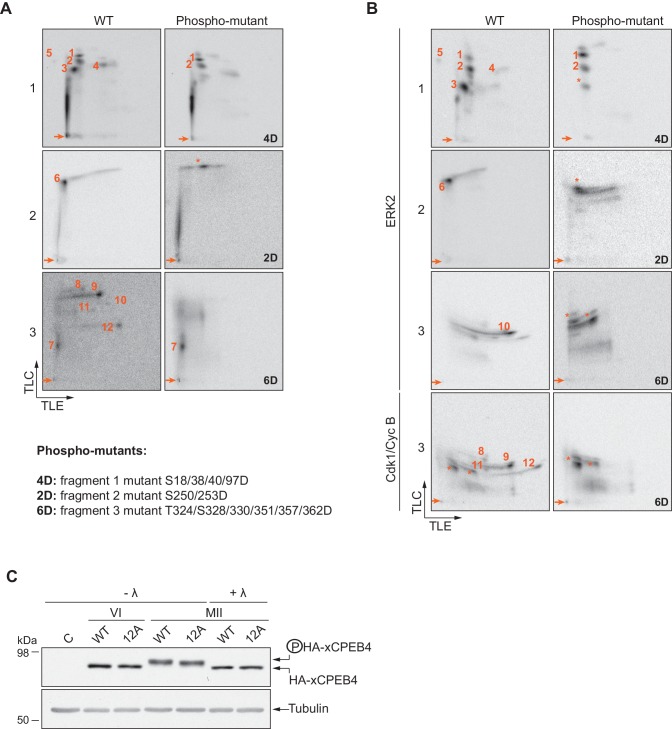
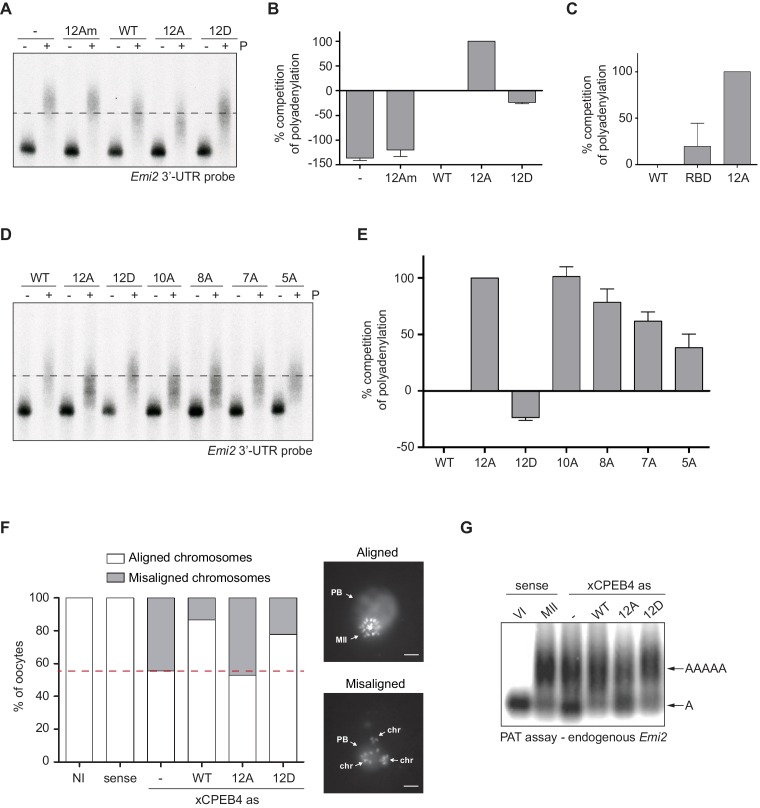
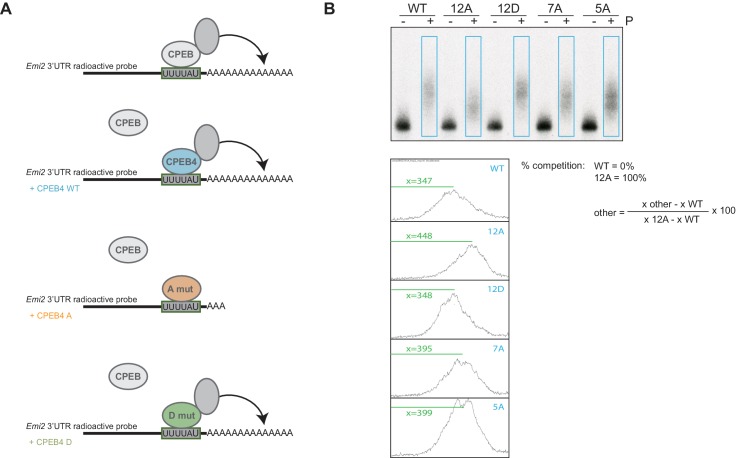
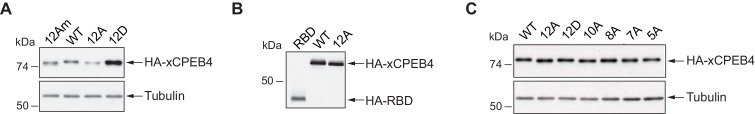
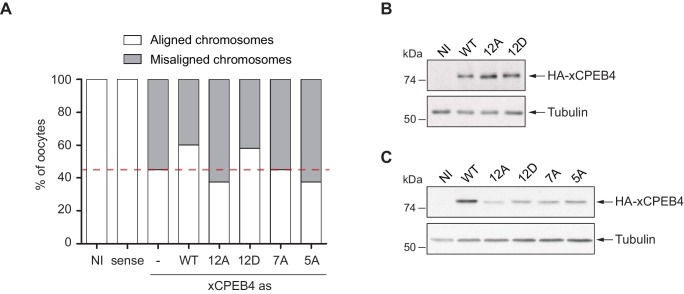
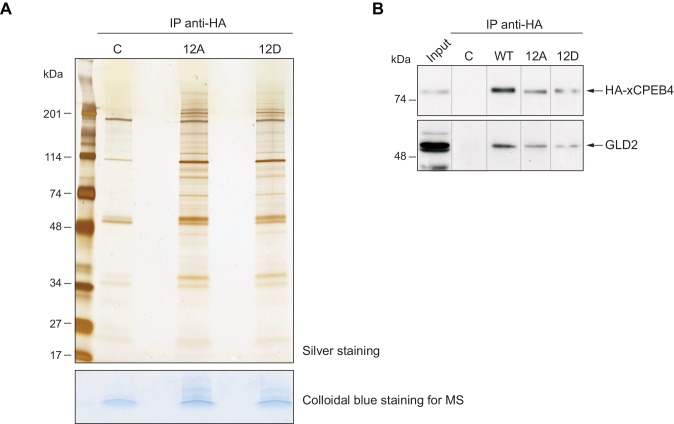
Guillén-Boixet Jordina, Buzon Víctor, Salvatella Xavier, Méndez Raúl
Institute for Research in Biomedicine, Barcelona, Spain.
The Barcelona Institute of Science and Technology, Barcelona, Spain.
Elife. 2016 Nov 1;5:e19298. doi: 10.7554/eLife.19298.
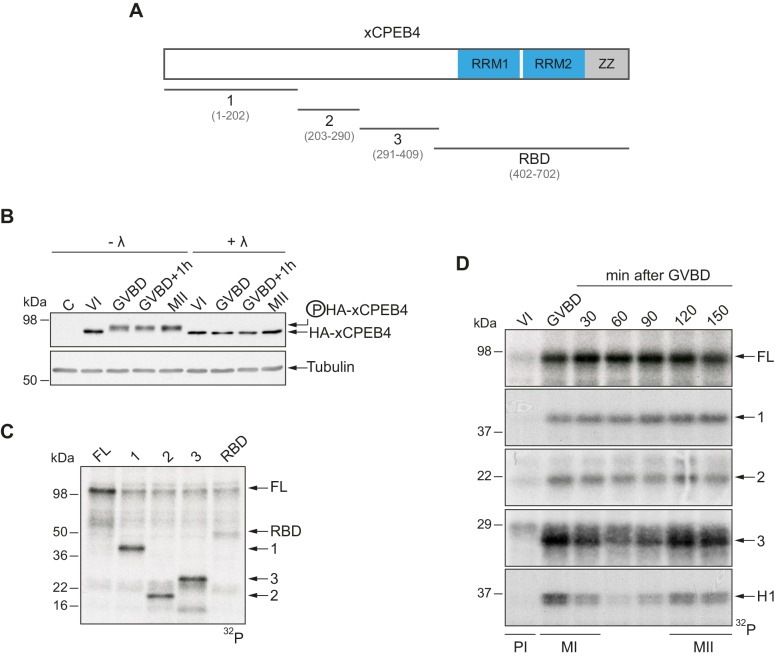
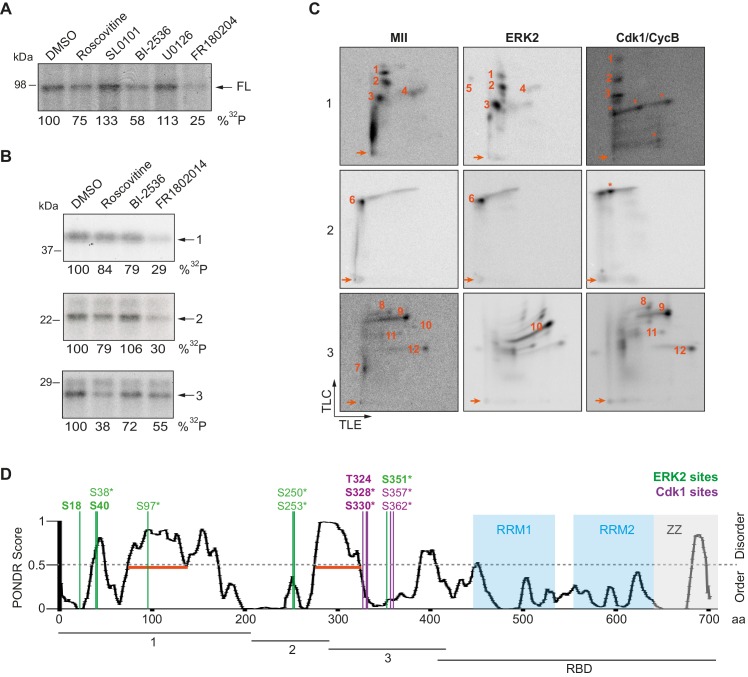
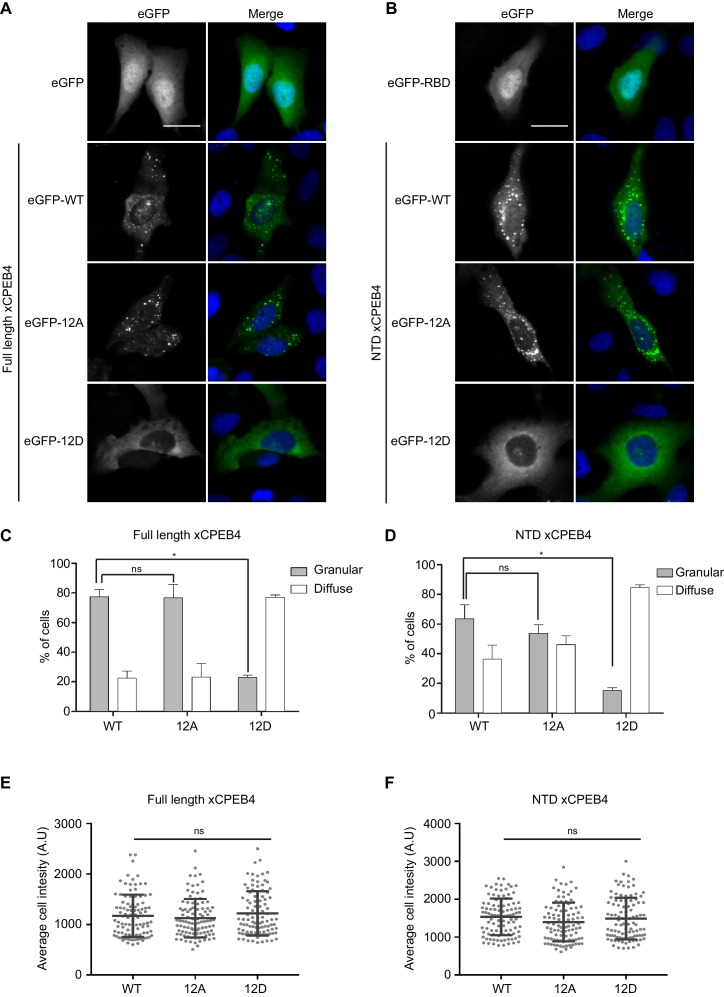
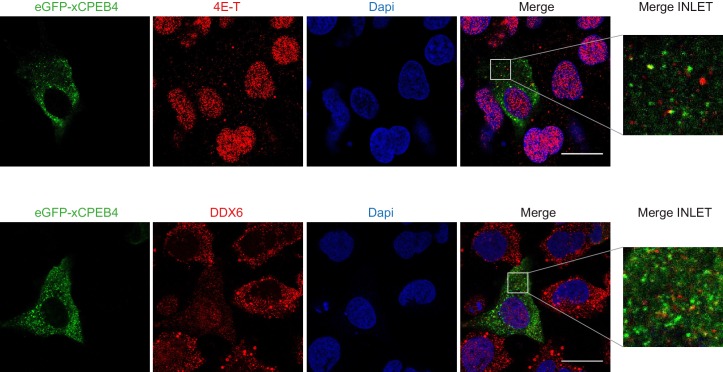
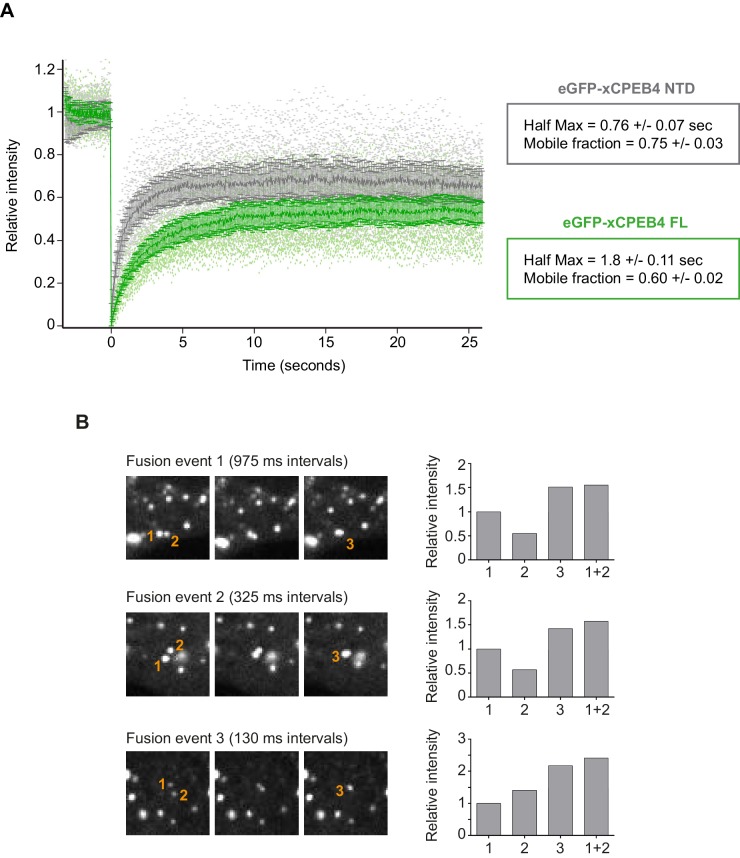
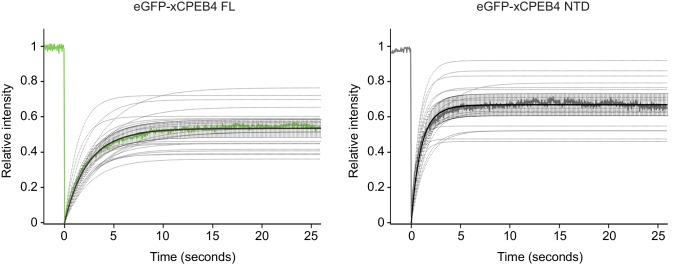
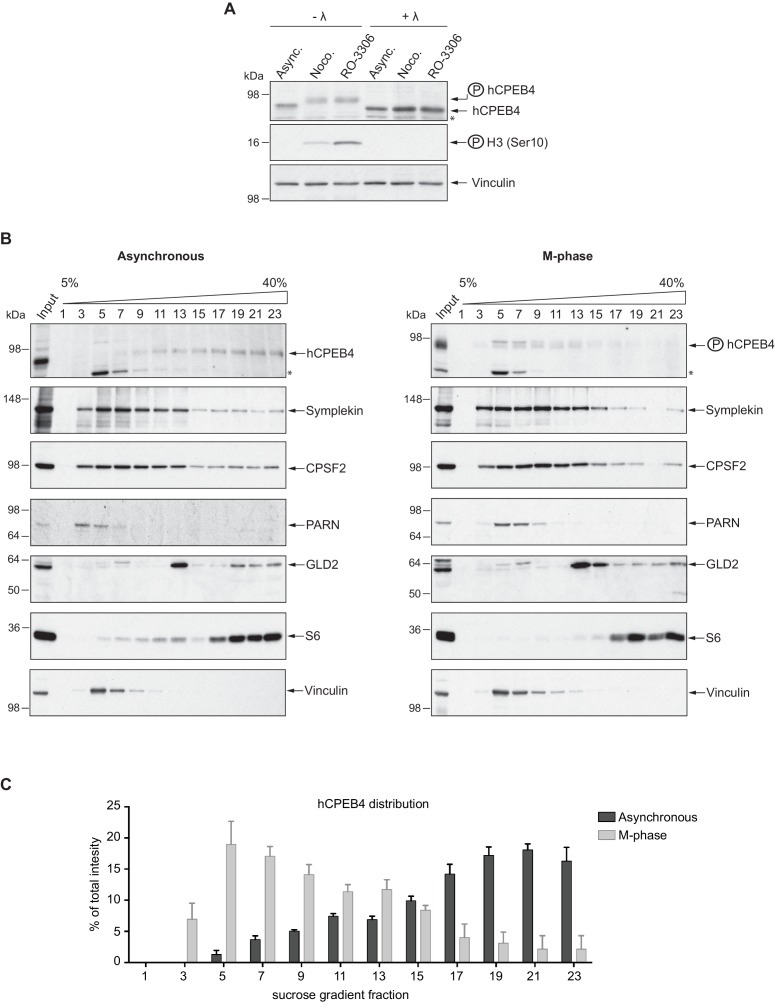
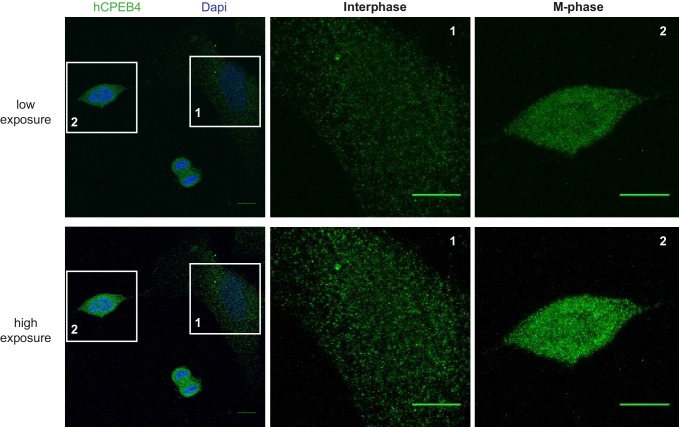
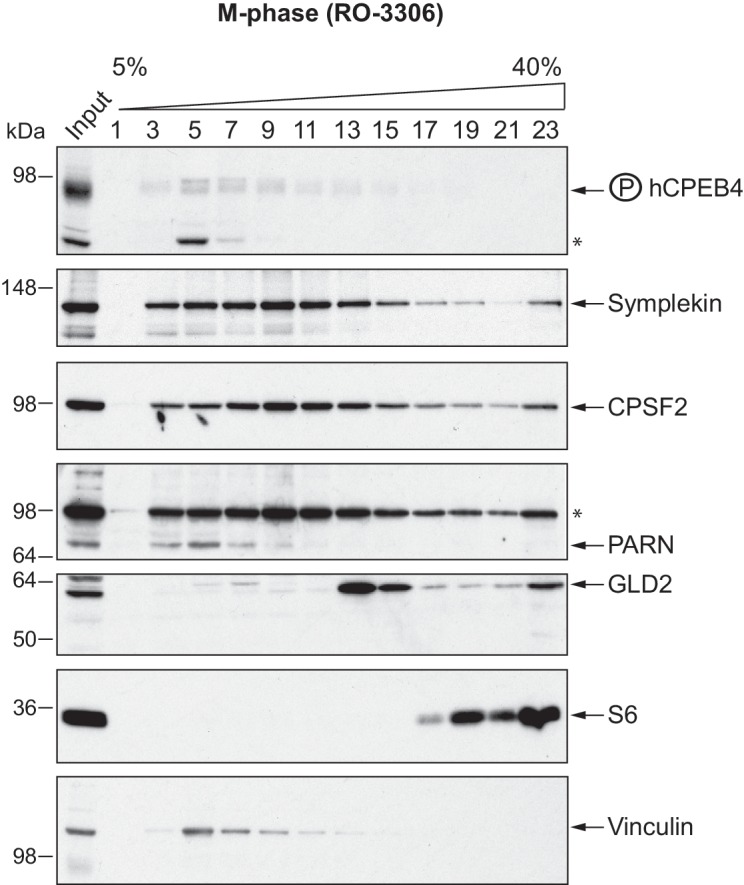
The four members of the vertebrate CPEB family of RNA-binding proteins share similar RNA-binding domains by which they regulate the translation of CPE-containing mRNAs, thereby controlling cell cycle and differentiation or synaptic plasticity. However, the N-terminal domains of CPEBs are distinct and contain specific regulatory post-translational modifications that presumably differentially integrate extracellular signals. Here we show that CPEB4 activity is regulated by ERK2- and Cdk1-mediated hyperphosphorylation. These phosphorylation events additively activate CPEB4 in M-phase by maintaining it in its monomeric state. In contrast, unphosphorylated CPEB4 phase separates into inactive, liquid-like droplets through its intrinsically disordered regions in the N-terminal domain. This dynamic and reversible regulation of CPEB4 is coordinated with that of CPEB1 through Cdk1, which inactivates CPEB1 while activating CPEB4, thereby integrating phase-specific signal transduction pathways to regulate cell cycle progression.
脊椎动物RNA结合蛋白CPEB家族的四个成员具有相似的RNA结合结构域,通过这些结构域它们调节含CPE的mRNA的翻译,从而控制细胞周期、分化或突触可塑性。然而,CPEB的N端结构域是不同的,并且包含特定的调节性翻译后修饰,这些修饰可能以不同方式整合细胞外信号。在此我们表明,CPEB4的活性受ERK2和Cdk1介导的过度磷酸化调节。这些磷酸化事件通过将CPEB4维持在单体状态,在M期累加激活CPEB4。相反,未磷酸化的CPEB4通过其N端结构域中固有无序区域相分离成无活性的液滴状。CPEB4这种动态且可逆的调节通过Cdk1与CPEB1的调节相协调,Cdk1使CPEB1失活同时激活CPEB4,从而整合阶段特异性信号转导途径以调节细胞周期进程。