Taylor Martin R G, Špírek Mário, Jian Ma Chu, Carzaniga Raffaella, Takaki Tohru, Collinson Lucy M, Greene Eric C, Krejci Lumir, Boulton Simon J
Clare Hall Laboratory, The Francis Crick Institute, South Mimms EN6 3LD, UK.
Department of Biology, Masaryk University, 62500 Brno, Czech Republic; International Clinical Research Center, St. Anne's University Hospital in Brno, 62500 Brno, Czech Republic.
Mol Cell. 2016 Dec 1;64(5):926-939. doi: 10.1016/j.molcel.2016.10.020. Epub 2016 Nov 17.
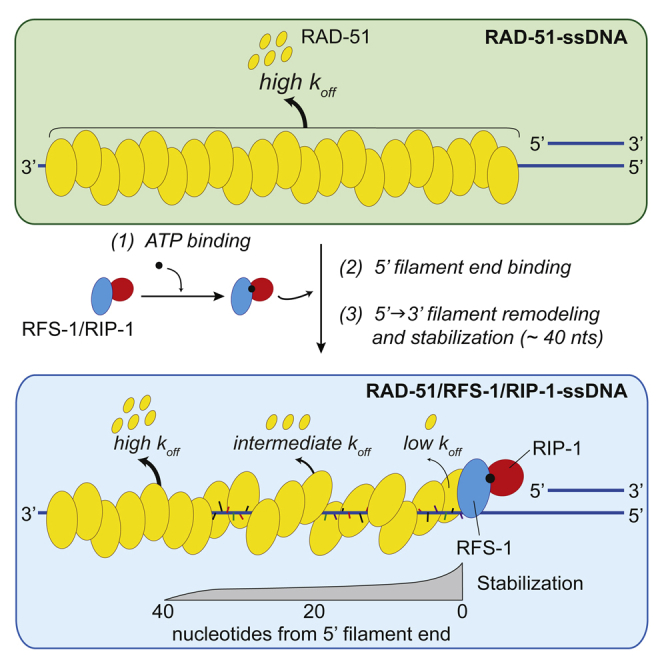
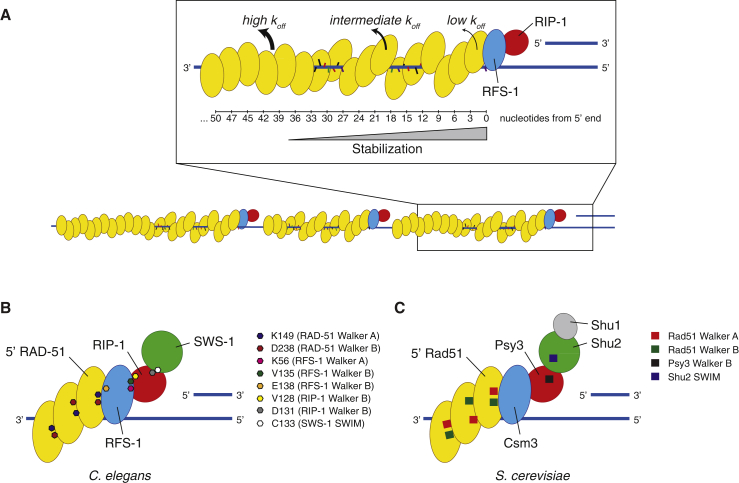
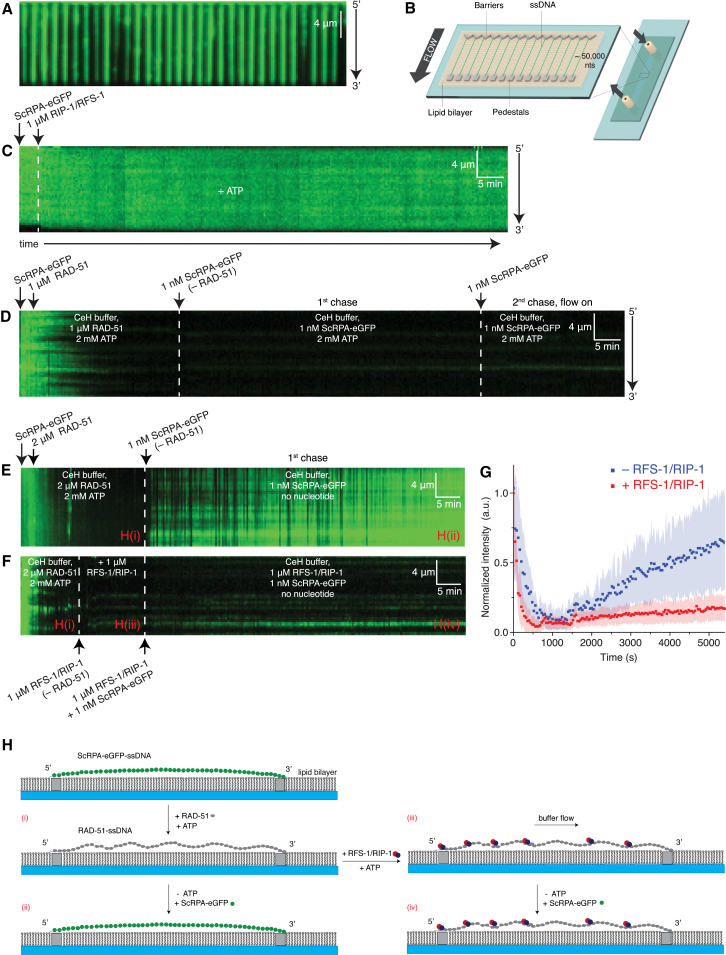
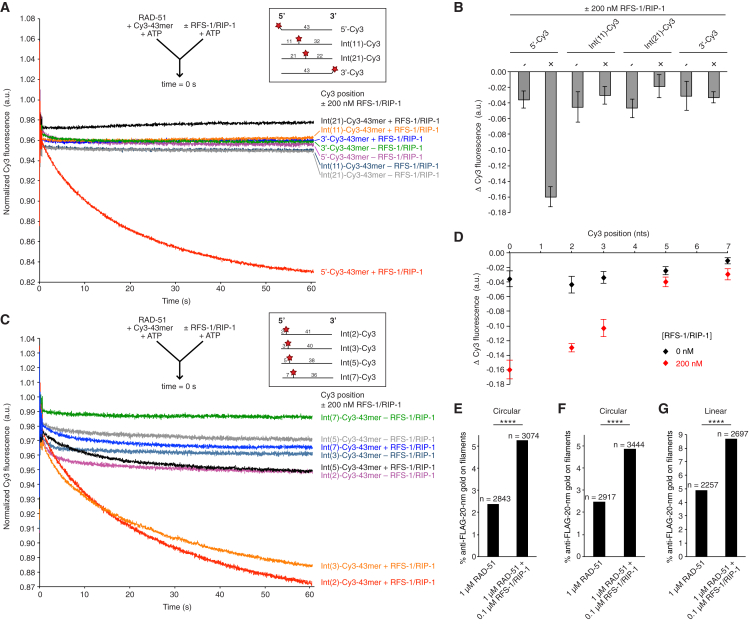
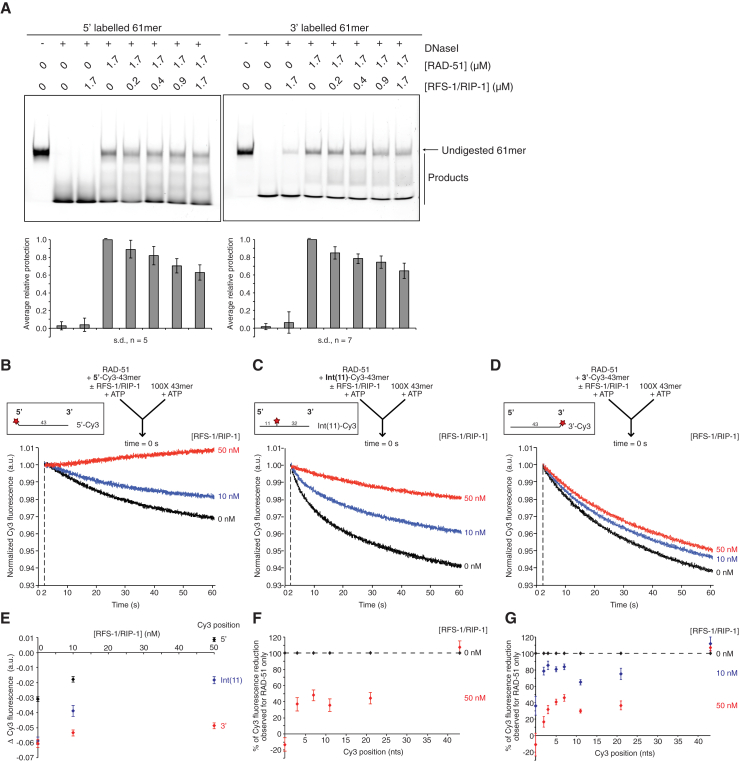
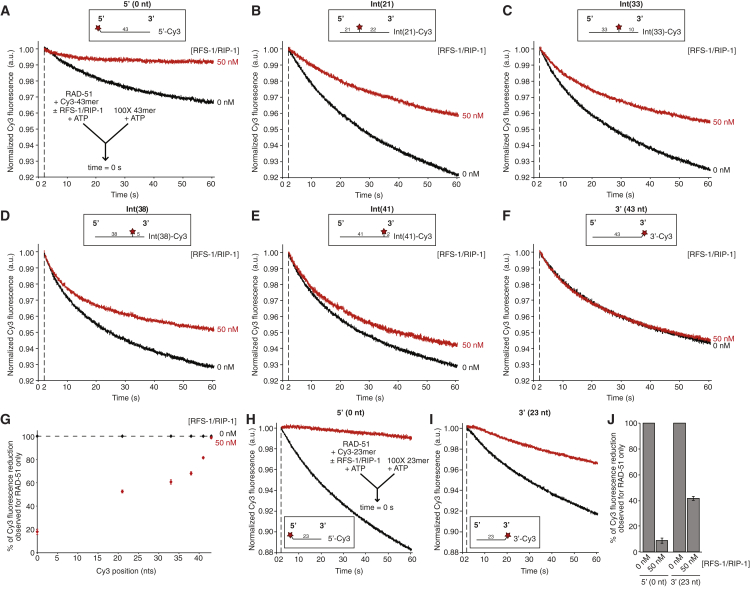
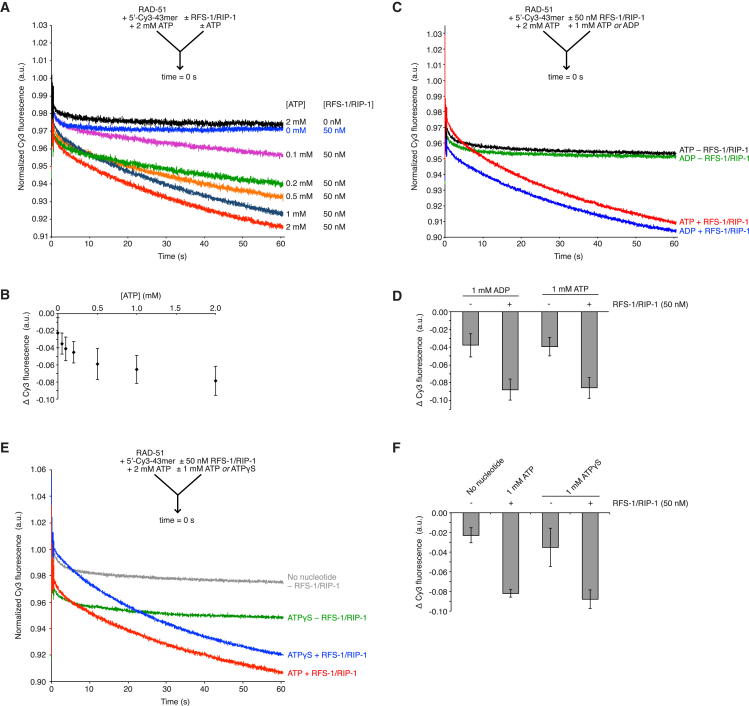
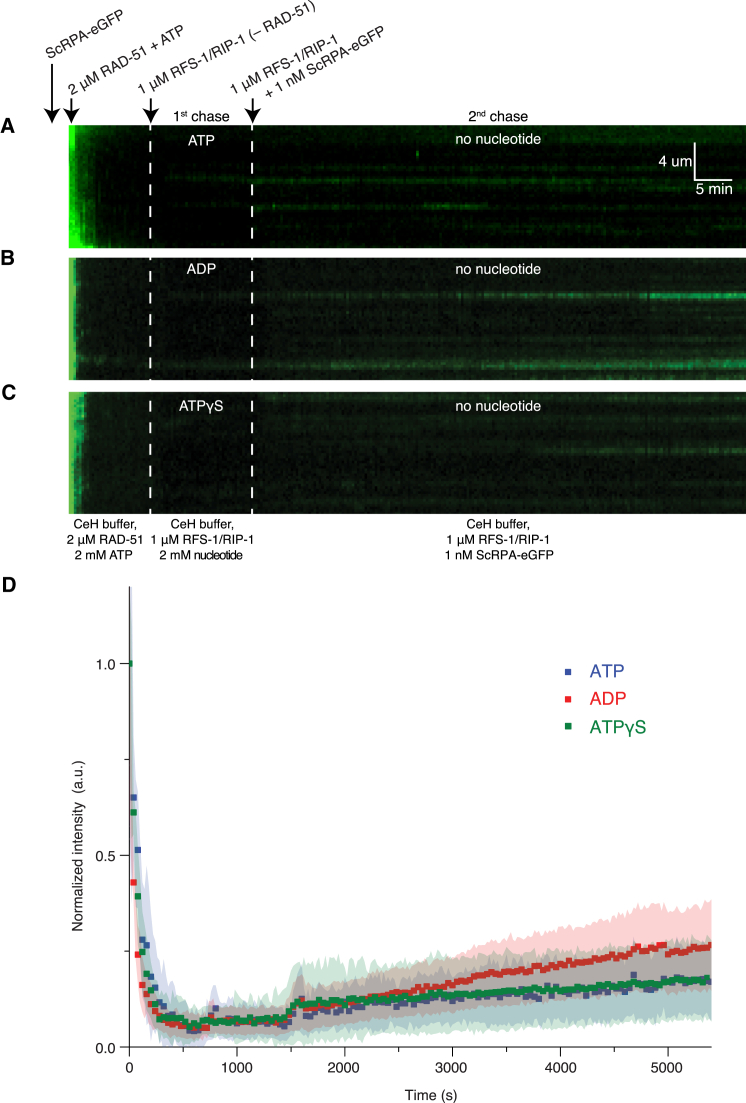
Central to homologous recombination in eukaryotes is the RAD51 recombinase, which forms helical nucleoprotein filaments on single-stranded DNA (ssDNA) and catalyzes strand invasion with homologous duplex DNA. Various regulatory proteins assist this reaction including the RAD51 paralogs. We recently discovered that a RAD51 paralog complex from C. elegans, RFS-1/RIP-1, functions predominantly downstream of filament assembly by binding and remodeling RAD-51-ssDNA filaments to a conformation more proficient for strand exchange. Here, we demonstrate that RFS-1/RIP-1 acts by shutting down RAD-51 dissociation from ssDNA. Using stopped-flow experiments, we show that RFS-1/RIP-1 confers this dramatic stabilization by capping the 5' end of RAD-51-ssDNA filaments. Filament end capping propagates a stabilizing effect with a 5'→3' polarity approximately 40 nucleotides along individual filaments. Finally, we discover that filament capping and stabilization are dependent on nucleotide binding, but not hydrolysis by RFS-1/RIP-1. These data define the mechanism of RAD51 filament remodeling by RAD51 paralogs.
真核生物同源重组的核心是RAD51重组酶,它在单链DNA(ssDNA)上形成螺旋核蛋白细丝,并催化与同源双链DNA的链入侵。各种调节蛋白协助这一反应,包括RAD51旁系同源物。我们最近发现,来自秀丽隐杆线虫的RAD51旁系同源物复合物RFS-1/RIP-1主要在细丝组装的下游发挥作用,通过结合并将RAD-51-ssDNA细丝重塑为更有利于链交换的构象。在这里,我们证明RFS-1/RIP-1通过阻止RAD-51从ssDNA上解离来发挥作用。通过停流实验,我们表明RFS-1/RIP-1通过封闭RAD-51-ssDNA细丝的5'端赋予这种显著的稳定性。细丝末端封闭沿着单个细丝以5'→3'极性传播约40个核苷酸的稳定作用。最后,我们发现细丝封闭和稳定依赖于核苷酸结合,但不依赖于RFS-1/RIP-1的水解。这些数据定义了RAD51旁系同源物对RAD51细丝重塑的机制。