Mowat Garth, Curtis P Jeff, Lafferty Diana J R
Natural Resource Science Section, BC Ministry of Forests, Lands and Natural Resource Operations, Nelson, British Columbia, Canada.
Department of Earth and Environmental Sciences, The University of British Columbia Okanagan Campus, Kelowna, British Columbia, Canada.
PLoS One. 2017 Mar 1;12(3):e0172194. doi: 10.1371/journal.pone.0172194. eCollection 2017.
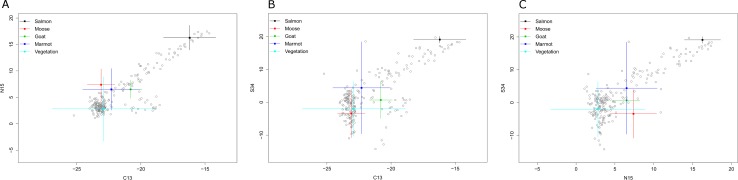
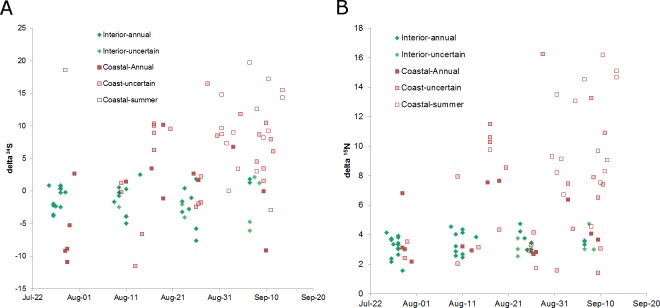
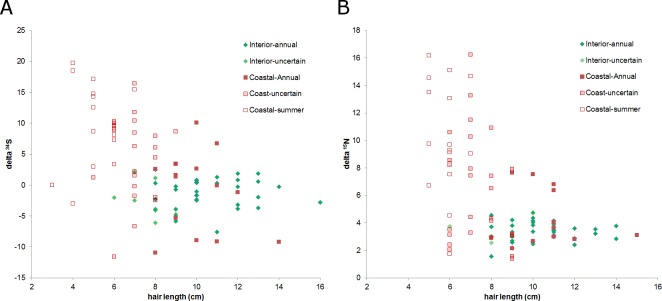
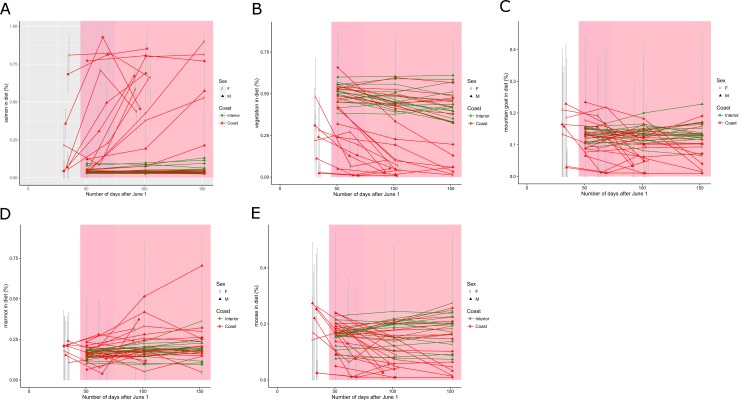
Stable isotope ratios of grizzly bear (Ursus arctos) guard hair collected from bears on the lower Stikine River, British Columbia (BC) were analyzed to: 1) test whether measuring δ34S values improved the precision of the salmon (Oncorhynchus spp.) diet fraction estimate relative to δ15N as is conventionally done, 2) investigate whether measuring δ34S values improves the separation of diet contributions of moose (Alces alces), marmot (Marmota caligata), and mountain goat (Oreamnos americanus) and, 3) examine the relationship between collection date and length of hair and stable isotope values. Variation in isotope signatures among hair samples from the same bear and year were not trivial. The addition of δ34S values to mixing models used to estimate diet fractions generated small improvement in the precision of salmon and terrestrial prey diet fractions. Although the δ34S value for salmon is precise and appears general among species and areas, sulfur ratios were strongly correlated with nitrogen ratios and therefore added little new information to the mixing model regarding the consumption of salmon. Mean δ34S values for the three terrestrial herbivores of interest were similar and imprecise, so these data also added little new information to the mixing model. The addition of sulfur data did confirm that at least some bears in this system ate marmots during summer and fall. We show that there are bears with short hair that assimilate >20% salmon in their diet and bears with longer hair that eat no salmon living within a few kilometers of one another in a coastal ecosystem. Grizzly bears are thought to re-grow hair between June and October however our analysis of sectioned hair suggested at least some hairs begin growing in July or August, not June and, that hair of wild bears may grow faster than observed in captive bears. Our hair samples may have been from the year of sampling or the previous year because samples were collected in summer when bears were growing new hair. The salmon diet fraction increased with later hair collection dates, as expected if samples were from the year of sampling because salmon began to arrive in mid-summer. Bears that ate salmon had shorter hair and δ15N and δ34S values declined with hair length, also suggesting some hair samples were grown the year of sampling. To be sure to capture an entire hair growth period, samples must be collected in late fall. Early spring samples are also likely to be from the previous year but the date when hair begins to grow appears to vary. Choosing the longest hair available should increase the chance the hair was grown during the previous year and, maximize the period for which diet is measured.
对从加拿大不列颠哥伦比亚省(BC)斯提金河下游的灰熊(棕熊,学名:Ursus arctos)身上采集的守护毛发的稳定同位素比率进行了分析,目的如下:1)测试相对于传统上使用的δ15N,测量δ34S值是否能提高鲑鱼(学名:Oncorhynchus spp.)饮食比例估计的精度;2)研究测量δ34S值是否能改善驼鹿(学名:Alces alces)、旱獭(学名:Marmota caligata)和北美野山羊(学名:Oreamnos americanus)饮食贡献的区分;3)研究采集日期与毛发长度及稳定同位素值之间的关系。来自同一只熊且年份相同的毛发样本间的同位素特征变化并非微不足道。在用于估计饮食比例的混合模型中加入δ34S值,对鲑鱼和陆地猎物饮食比例估计精度的提升幅度较小。尽管鲑鱼的δ34S值较为精确,且在不同物种和区域似乎具有普遍性,但硫比率与氮比率强烈相关,因此在混合模型中关于鲑鱼消费方面几乎没有增加新信息。三种目标陆地食草动物的平均δ34S值相似且不精确,所以这些数据在混合模型中也几乎没有增加新信息。硫数据的加入确实证实,该系统中至少有一些熊在夏季和秋季食用旱獭。我们发现,在一个沿海生态系统中,相距几公里内存在毛发短且饮食中鲑鱼同化量超过20%的熊,以及毛发长且不吃鲑鱼的熊。灰熊被认为在6月至10月之间重新生长毛发,然而我们对分段毛发的分析表明,至少一些毛发在7月或8月开始生长,而非6月,并且野生熊的毛发可能比圈养熊的生长速度更快。我们的毛发样本可能来自采样当年或前一年,因为样本是在夏季熊生长新毛发时采集的。如预期的那样,如果样本来自采样当年,鲑鱼饮食比例会随着毛发采集日期的推迟而增加,因为鲑鱼在仲夏开始抵达。食用鲑鱼的熊毛发较短,δ15N和δ34S值随毛发长度下降,这也表明一些毛发样本是在采样当年生长的。为确保捕捉到整个毛发生长期,样本必须在深秋采集。早春样本也可能来自前一年,但毛发开始生长的日期似乎有所不同。选择最长的可用毛发应能增加毛发在前一年生长的几率,并最大化测量饮食的时间段。