Sebro Ronnie, Peloso Gina M, Dupuis Josée, Risch Neil J
Department of Radiology, University of Pennsylvania, Philadelphia, PA, United States of America.
Department of Biostatistics, Boston University School of Public Health, Boston, MA, United States of America.
PLoS Genet. 2017 Apr 6;13(4):e1006655. doi: 10.1371/journal.pgen.1006655. eCollection 2017 Apr.
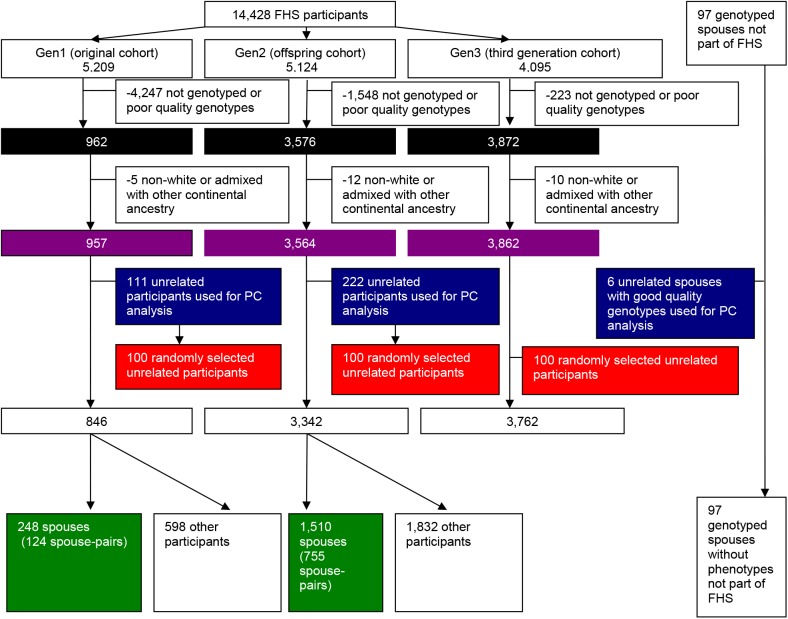
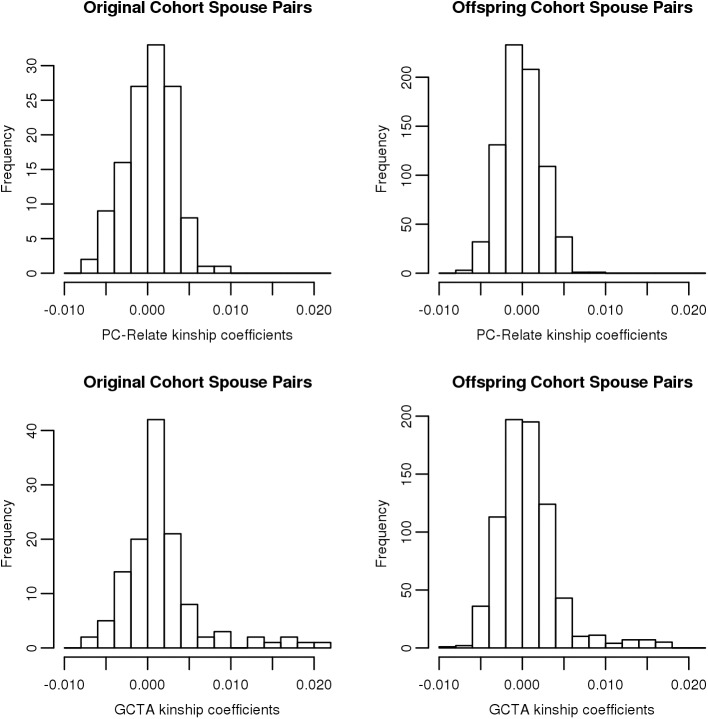
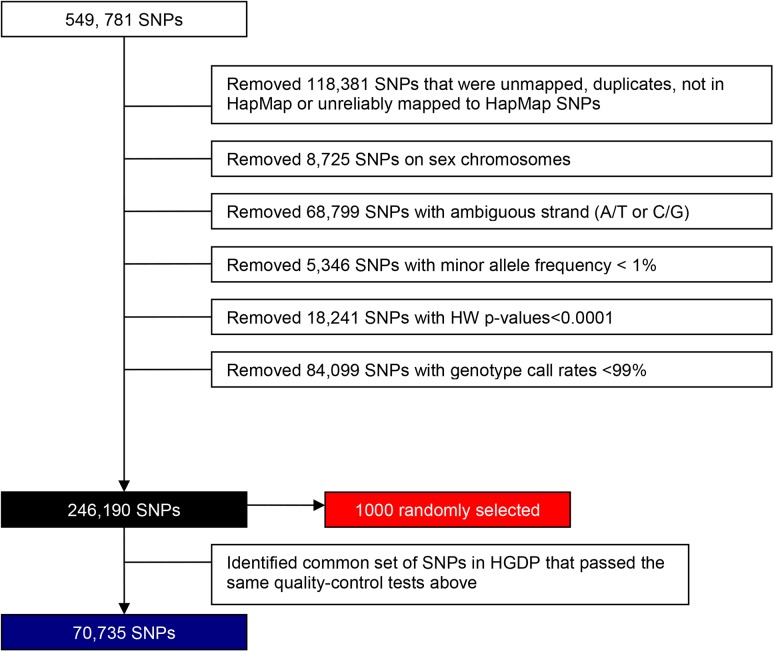
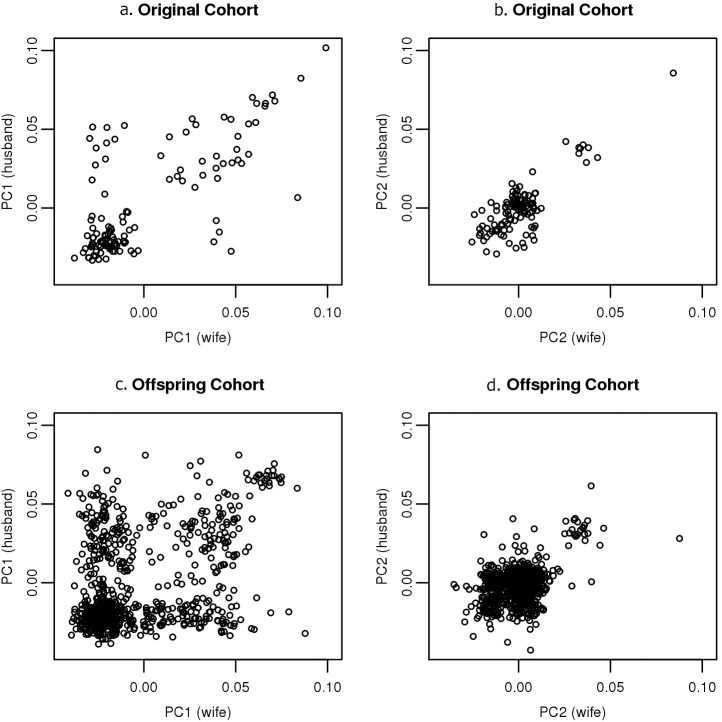
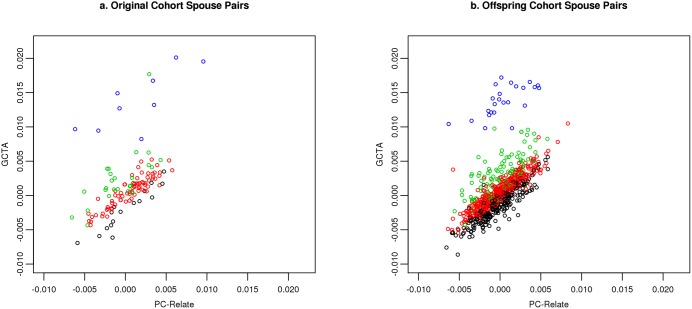
Genetic similarity of spouses can reflect factors influencing mate choice, such as physical/behavioral characteristics, and patterns of social endogamy. Spouse correlations for both genetic ancestry and measured traits may impact genotype distributions (Hardy Weinberg and linkage equilibrium), and therefore genetic association studies. Here we evaluate white spouse-pairs from the Framingham Heart Study (FHS) original and offspring cohorts (N = 124 and 755, respectively) to explore spousal genetic similarity and its consequences. Two principal components (PCs) of the genome-wide association (GWA) data were identified, with the first (PC1) delineating clines of Northern/Western to Southern European ancestry and the second (PC2) delineating clines of Ashkenazi Jewish ancestry. In the original (older) cohort, there was a striking positive correlation between the spouses in PC1 (r = 0.73, P = 3x10(-22)) and also for PC2 (r = 0.80, P = 7x10(-29)). In the offspring cohort, the spouse correlations were lower but still highly significant for PC1 (r = 0.38, P = 7x10(-28)) and for PC2 (r = 0.45, P = 2x10(-39)). We observed significant Hardy-Weinberg disequilibrium for single nucleotide polymorphisms (SNPs) loading heavily on PC1 and PC2 across 3 generations, and also significant linkage disequilibrium between unlinked SNPs; both decreased with time, consistent with reduced ancestral endogamy over generations and congruent with theoretical calculations. Ignoring ancestry, estimates of spouse kinship have a mean significantly greater than 0, and more so in the earlier generations. Adjusting kinship estimates for genetic ancestry through the use of PCs led to a mean spouse kinship not different from 0, demonstrating that spouse genetic similarity could be fully attributed to ancestral assortative mating. These findings also have significance for studies of heritability that are based on distantly related individuals (kinship less than 0.05), as we also demonstrate the poor correlation of kinship estimates in that range when ancestry is or is not taken into account.
配偶的基因相似性能够反映影响配偶选择的因素,如身体/行为特征以及社会内婚模式。基因血统和测量性状的配偶相关性可能会影响基因型分布(哈迪-温伯格平衡和连锁平衡),进而影响基因关联研究。在此,我们评估了弗雷明汉心脏研究(FHS)原始队列和后代队列中的白人配偶对(分别为N = 124对和755对),以探究配偶间的基因相似性及其影响。我们确定了全基因组关联(GWA)数据的两个主要成分(PCs),第一个(PC1)描绘了北欧/西欧到南欧血统的渐变群,第二个(PC2)描绘了阿什肯纳兹犹太血统的渐变群。在原始(较年长)队列中,配偶在PC1上存在显著的正相关(r = 0.73,P = 3×10⁻²²),在PC2上也如此(r = 0.80,P = 7×10⁻²⁹)。在后代队列中,配偶相关性较低,但在PC1上仍高度显著(r = 0.38,P = 7×10⁻²⁸),在PC2上也是如此(r = 0.45,P = 2×10⁻³⁹)。我们观察到在三代中,在PC1和PC2上大量负荷的单核苷酸多态性(SNPs)存在显著的哈迪-温伯格不平衡,并且非连锁SNPs之间也存在显著的连锁不平衡;两者都随时间减少,这与几代人之间祖先内婚率降低一致,并且与理论计算相符。忽略血统,配偶亲属关系的估计均值显著大于0,在早期几代中更是如此。通过使用PCs对基因血统进行亲属关系估计调整后,配偶亲属关系的均值与0无异,这表明配偶的基因相似性可完全归因于祖先的选型交配。这些发现对于基于远亲个体(亲属关系小于0.05)的遗传力研究也具有重要意义,因为我们还证明了在考虑或不考虑血统时,该范围内亲属关系估计的相关性较差。