Di Giorgio Eros, Franforte Elisa, Cefalù Sebastiano, Rossi Sabrina, Dei Tos Angelo Paolo, Brenca Monica, Polano Maurizio, Maestro Roberta, Paluvai Harikrishnareddy, Picco Raffaella, Brancolini Claudio
Department of Medical and Biological Sciences, Università degli Studi di Udine. P.le Kolbe 4-Udine Italy.
Department of Anatomical Pathology, Treviso General Hospital, Treviso, Italy.
PLoS Genet. 2017 Apr 18;13(4):e1006752. doi: 10.1371/journal.pgen.1006752. eCollection 2017 Apr.
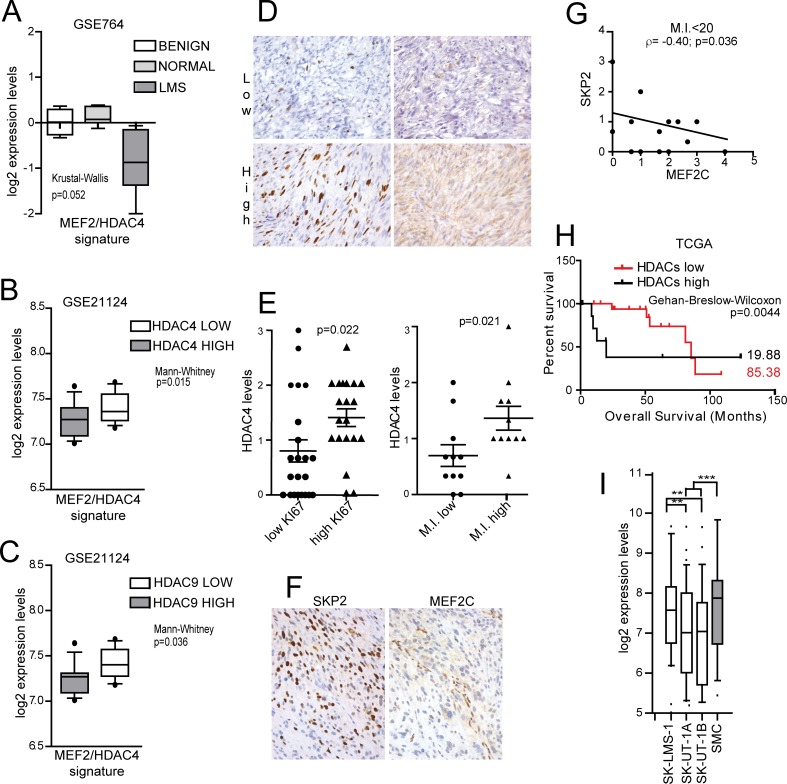
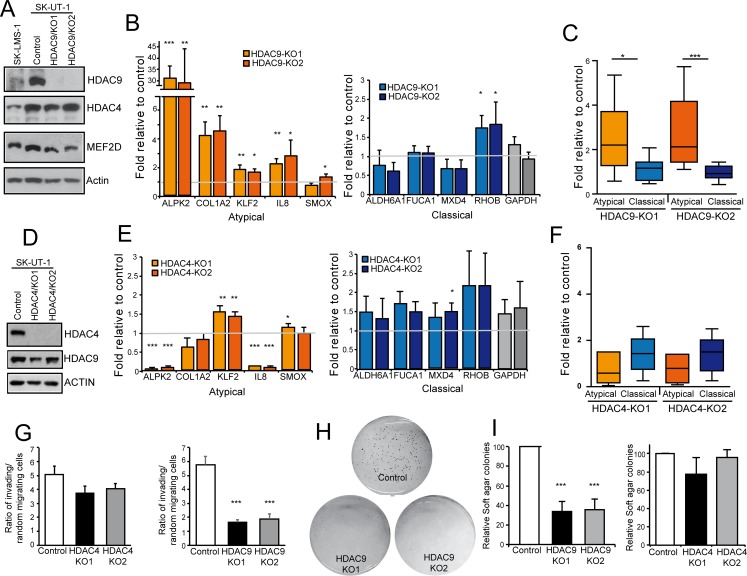
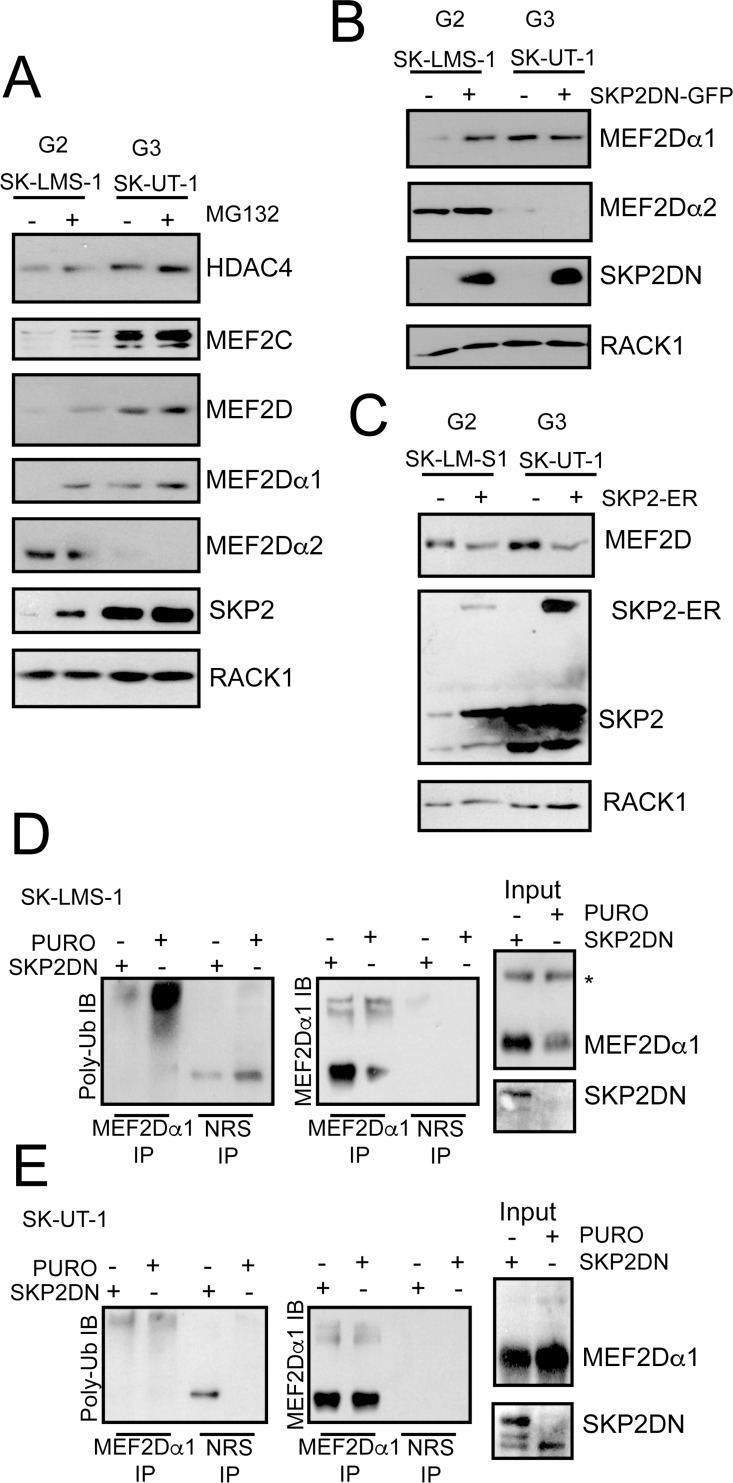
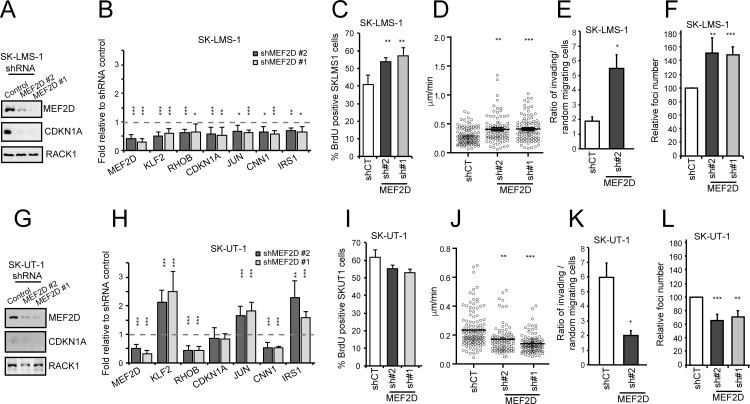
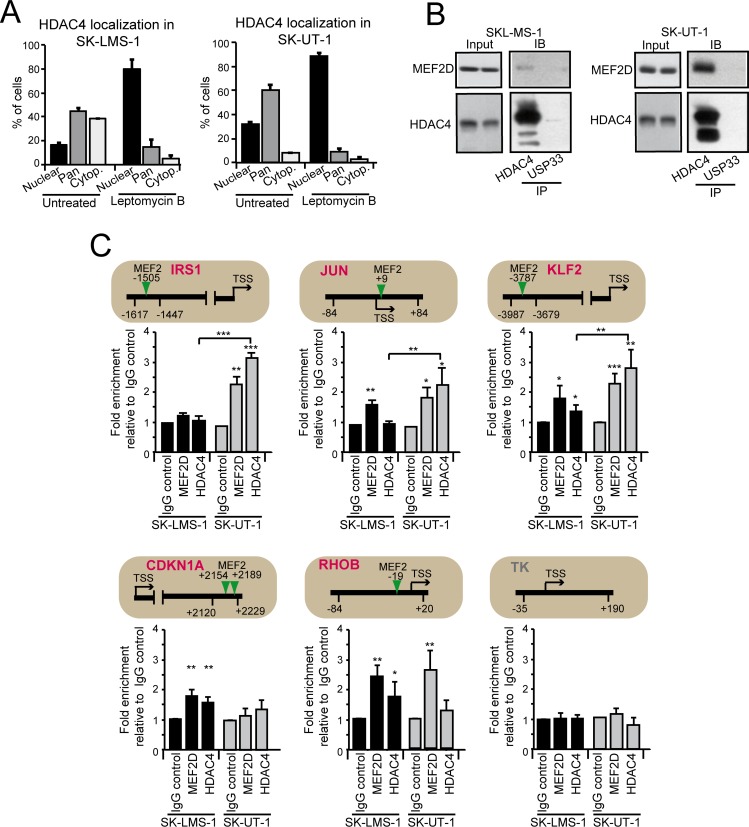
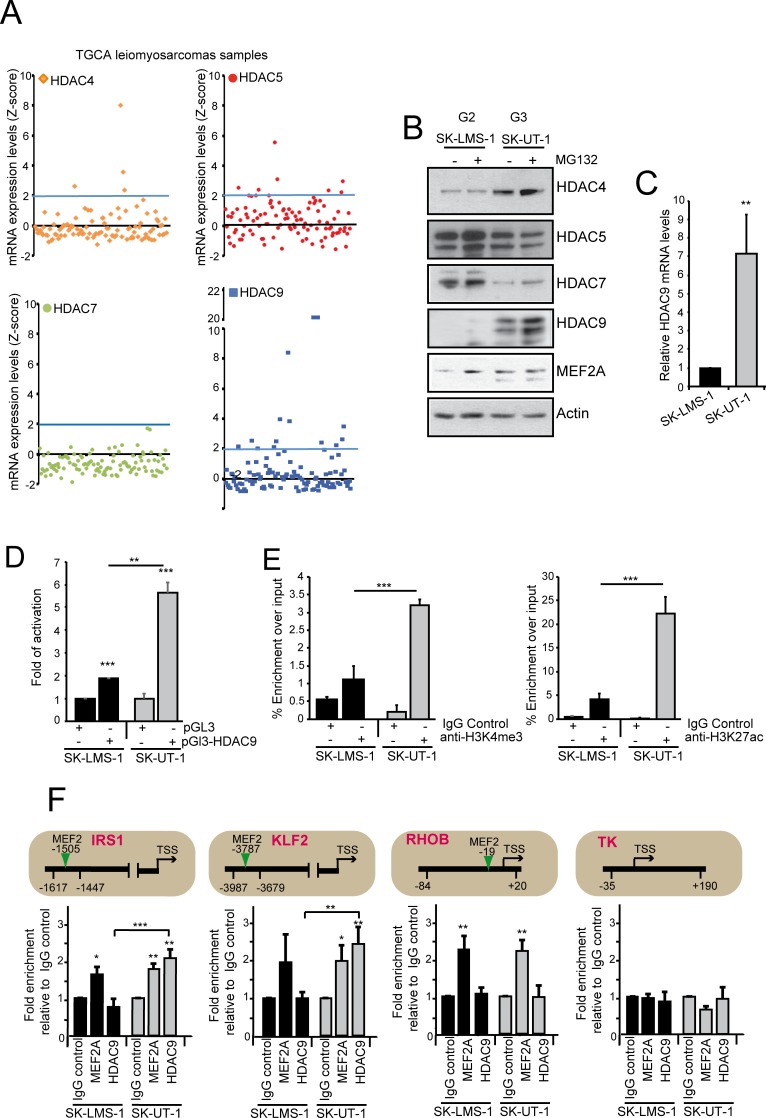
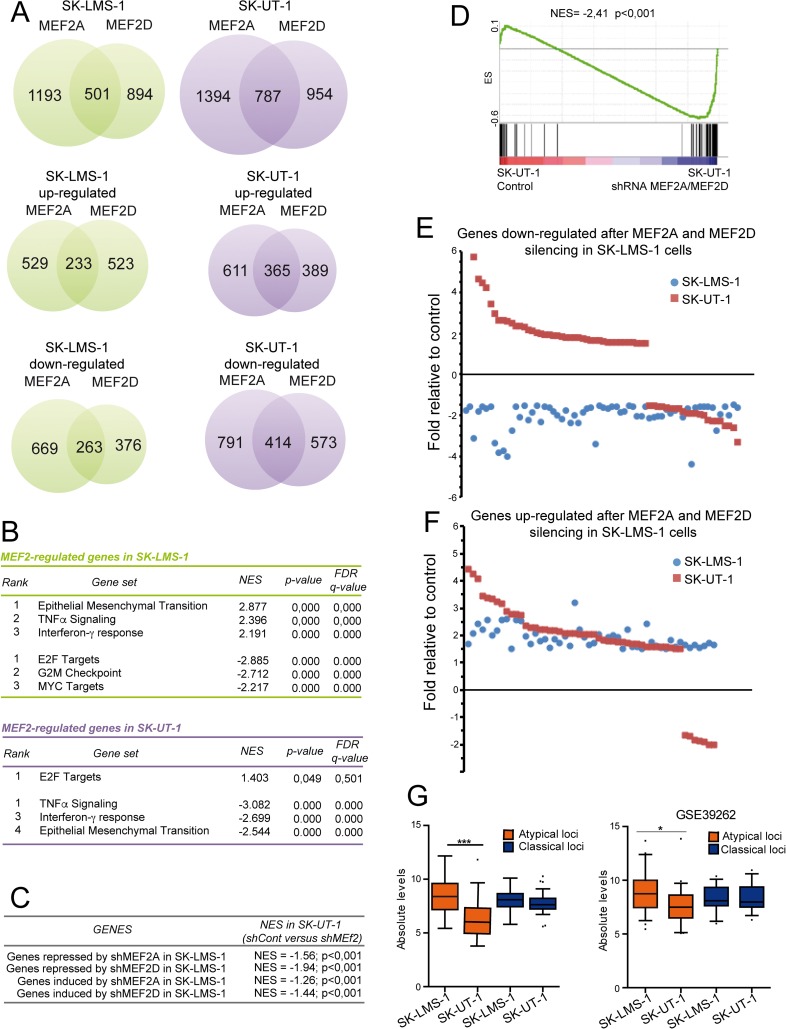
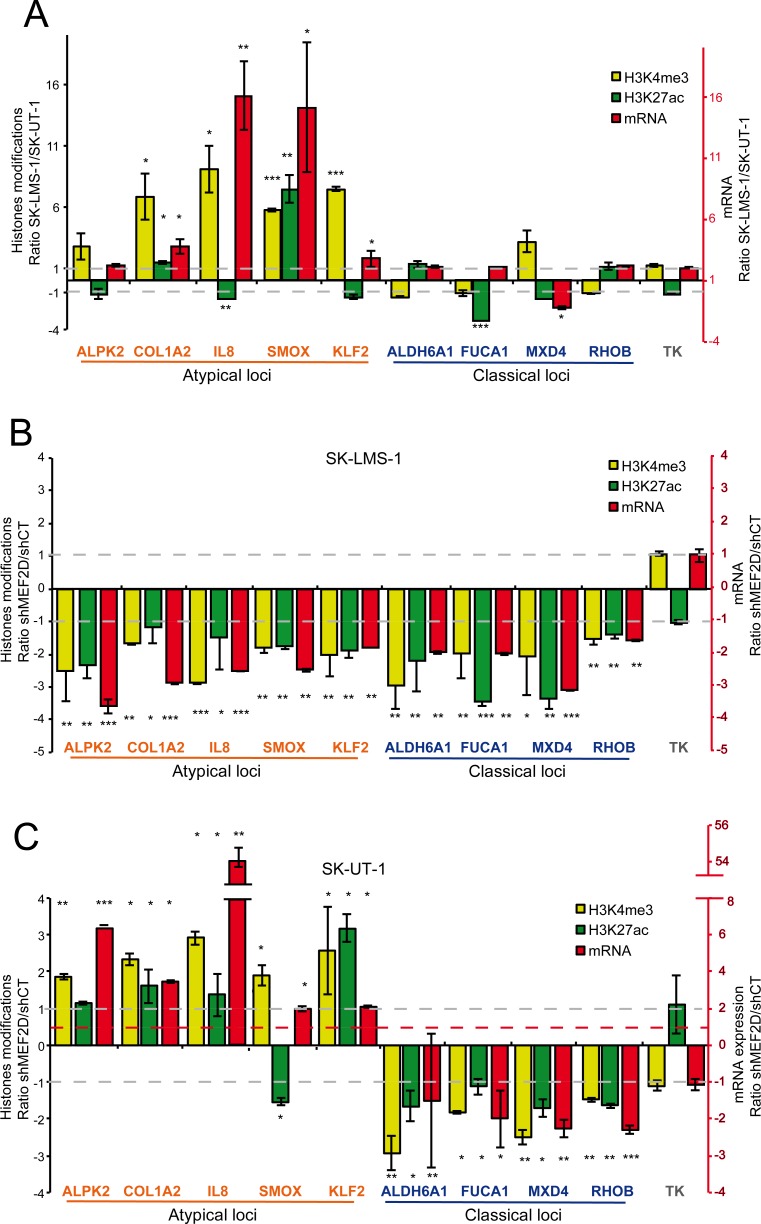
The contribution of MEF2 TFs to the tumorigenic process is still mysterious. Here we clarify that MEF2 can support both pro-oncogenic or tumor suppressive activities depending on the interaction with co-activators or co-repressors partners. Through these interactions MEF2 supervise histone modifications associated with gene activation/repression, such as H3K4 methylation and H3K27 acetylation. Critical switches for the generation of a MEF2 repressive environment are class IIa HDACs. In leiomyosarcomas (LMS), this two-faced trait of MEF2 is relevant for tumor aggressiveness. Class IIa HDACs are overexpressed in 22% of LMS, where high levels of MEF2, HDAC4 and HDAC9 inversely correlate with overall survival. The knock out of HDAC9 suppresses the transformed phenotype of LMS cells, by restoring the transcriptional proficiency of some MEF2-target loci. HDAC9 coordinates also the demethylation of H3K4me3 at the promoters of MEF2-target genes. Moreover, we show that class IIa HDACs do not bind all the regulative elements bound by MEF2. Hence, in a cell MEF2-target genes actively transcribed and strongly repressed can coexist. However, these repressed MEF2-targets are poised in terms of chromatin signature. Overall our results candidate class IIa HDACs and HDAC9 in particular, as druggable targets for a therapeutic intervention in LMS.
MEF2转录因子在肿瘤发生过程中的作用仍不清楚。在此我们阐明,MEF2根据与共激活因子或共抑制因子伙伴的相互作用,既能支持促癌活性,也能支持肿瘤抑制活性。通过这些相互作用,MEF2调控与基因激活/抑制相关的组蛋白修饰,如H3K4甲基化和H3K27乙酰化。产生MEF2抑制环境的关键开关是IIa类组蛋白去乙酰化酶(HDAC)。在平滑肌肉瘤(LMS)中,MEF2的这种两面性与肿瘤侵袭性相关。22%的LMS中IIa类HDAC过表达,其中MEF2、HDAC4和HDAC9的高水平与总生存期呈负相关。敲除HDAC9可恢复一些MEF2靶基因座的转录能力,从而抑制LMS细胞的转化表型。HDAC9还协调MEF2靶基因启动子处H3K4me3的去甲基化。此外,我们发现IIa类HDAC并不结合MEF2结合的所有调控元件。因此,在一个细胞中,被MEF2靶向的活跃转录和强烈抑制的基因可以共存。然而,这些被抑制的MEF2靶基因在染色质特征方面处于准备状态。总体而言,我们的结果表明IIa类HDAC,特别是HDAC9,可作为LMS治疗干预的可药物靶向靶点。