Department of Anatomy and Cell Biology, Oklahoma State University Center for Health Sciences, Tulsa, Oklahoma, 74107-1898, USA.
Department of Biological Science, Florida State University, Tallahassee, Florida, 32306-4295, USA.
Sci Rep. 2017 May 17;7(1):2012. doi: 10.1038/s41598-017-02161-w.
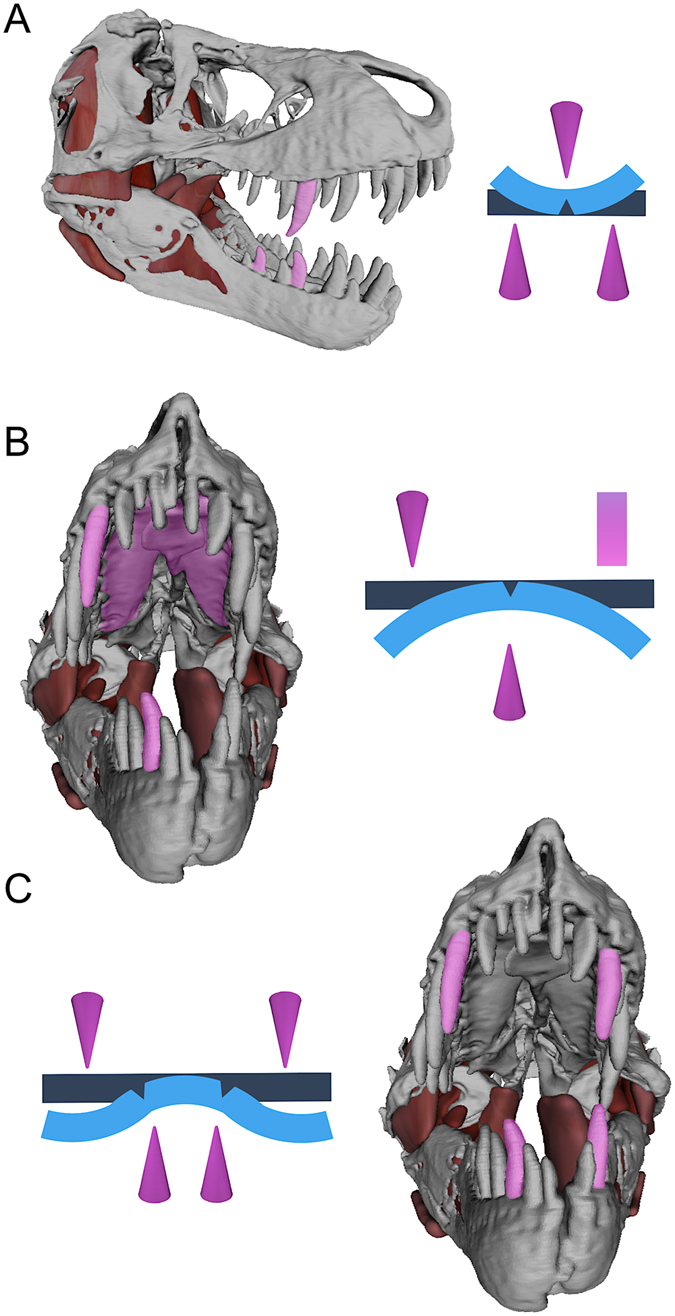
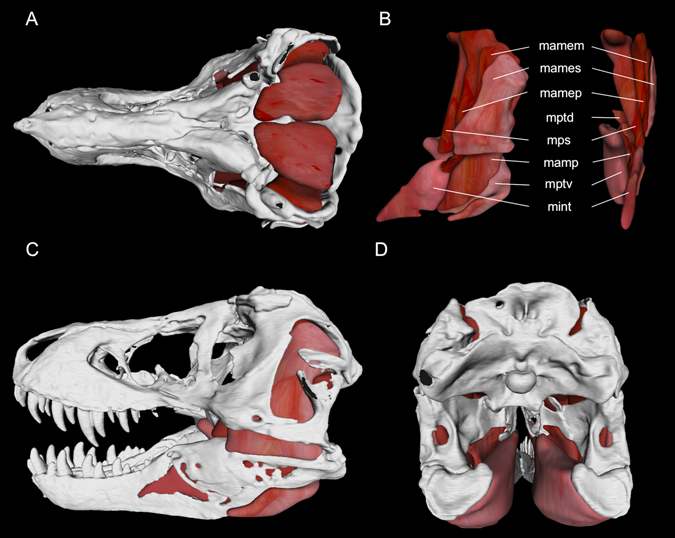
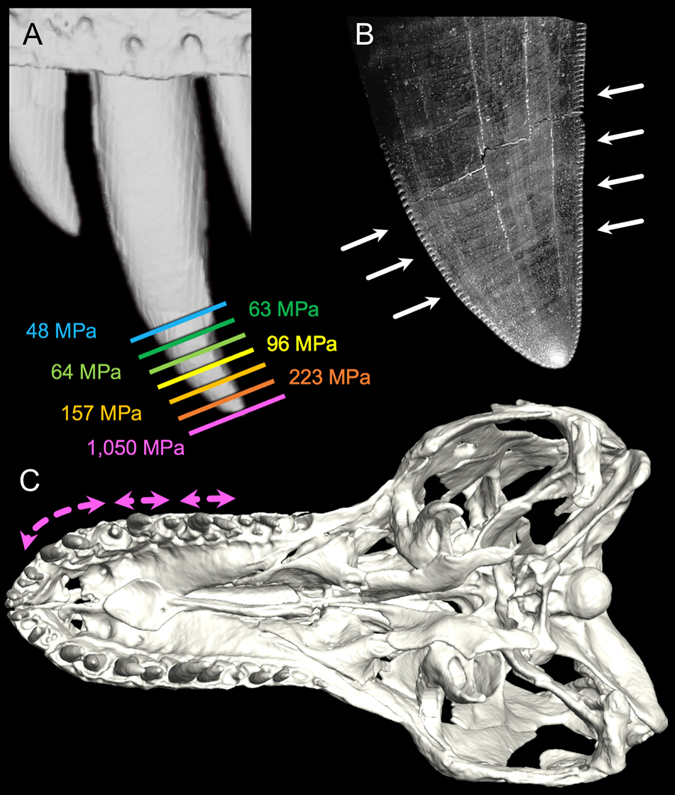
Most carnivorous mammals can pulverize skeletal elements by generating tooth pressures between occluding teeth that exceed cortical bone shear strength, thereby permitting access to marrow and phosphatic salts. Conversely, carnivorous reptiles have non-occluding dentitions that engender negligible bone damage during feeding. As a result, most reptilian predators can only consume bones in their entirety. Nevertheless, North American tyrannosaurids, including the giant (13 metres [m]) theropod dinosaur Tyrannosaurus rex stand out for habitually biting deeply into bones, pulverizing and digesting them. How this mammal-like capacity was possible, absent dental occlusion, is unknown. Here we analyzed T. rex feeding behaviour from trace evidence, estimated bite forces and tooth pressures, and studied tooth-bone contacts to provide the answer. We show that bone pulverization was made possible through a combination of: (1) prodigious bite forces (8,526-34,522 newtons [N]) and tooth pressures (718-2,974 megapascals [MPa]) promoting crack propagation in bones, (2) tooth form and dental arcade configurations that concentrated shear stresses, and (3) repetitive, localized biting. Collectively, these capacities and behaviors allowed T. rex to finely fragment bones and more fully exploit large dinosaur carcasses for sustenance relative to competing carnivores.
大多数肉食性哺乳动物可以通过在咬合牙齿之间产生超过皮质骨剪切强度的牙齿压力来粉碎骨骼元素,从而可以获得骨髓和磷酸盐盐。相反,肉食性爬行动物具有非咬合的牙齿,在进食过程中几乎不会对骨骼造成损伤。因此,大多数爬行动物捕食者只能完整地消耗骨头。然而,北美暴龙类,包括巨大的(13 米)兽脚亚目恐龙霸王龙,以习惯性地深入骨头、粉碎和消化骨头而著称。在没有牙齿咬合的情况下,这种类似哺乳动物的能力是如何实现的尚不清楚。在这里,我们从痕迹证据分析了霸王龙的进食行为,估计了咬合力和牙齿压力,并研究了牙齿与骨骼的接触情况,以提供答案。我们表明,骨粉碎是通过以下几种方式实现的:(1)巨大的咬合力(8526-34522 牛顿(N))和牙齿压力(718-2974 兆帕斯卡(MPa))促进了骨骼中的裂纹扩展,(2)牙齿形状和牙弓配置集中了剪切应力,(3)反复的、局部的咬伤。总的来说,这些能力和行为使霸王龙能够精细地粉碎骨头,并更充分地利用大型恐龙尸体来维持生存,相对于竞争的肉食动物。