Boronat Susanna, Domènech Alba, Carmona Mercè, García-Santamarina Sarela, Bañó M Carmen, Ayté José, Hidalgo Elena
Oxidative Stress and Cell Cycle Group, Universitat Pompeu Fabra, Barcelona, Spain.
Departamento de Bioquímica y Biología Molecular, Universitat de València, Valencia, Spain.
PLoS Genet. 2017 Jun 22;13(6):e1006858. doi: 10.1371/journal.pgen.1006858. eCollection 2017 Jun.
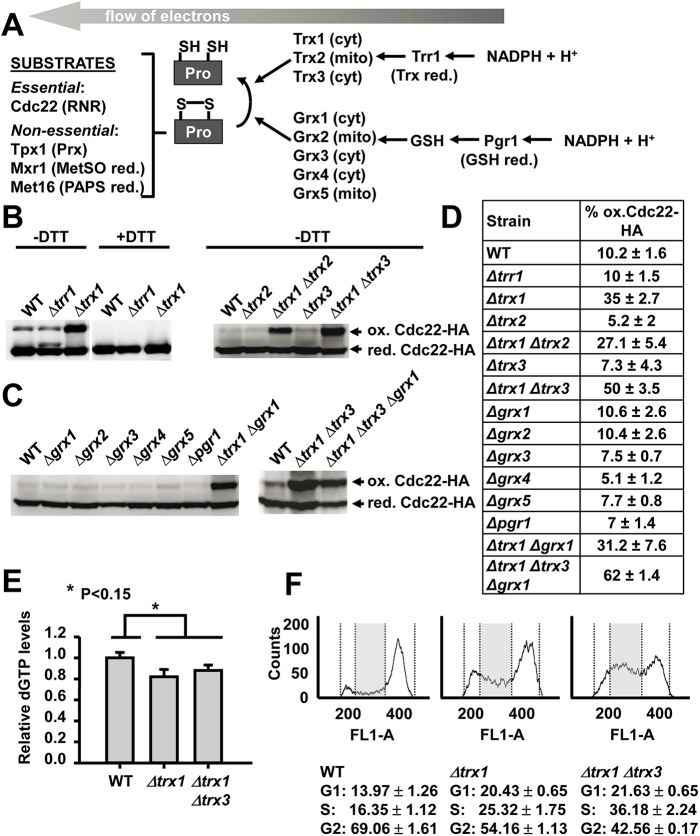
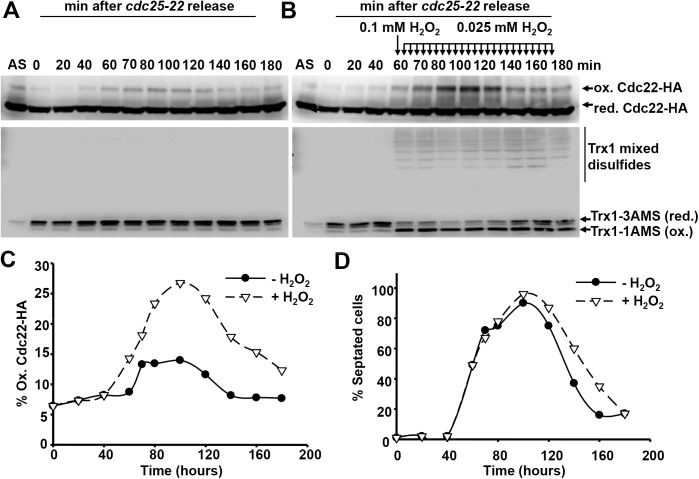
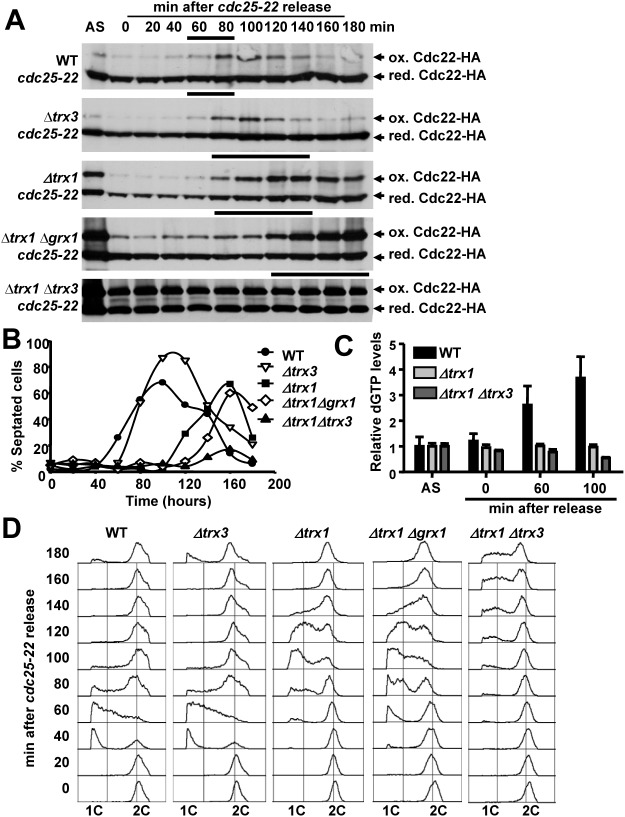
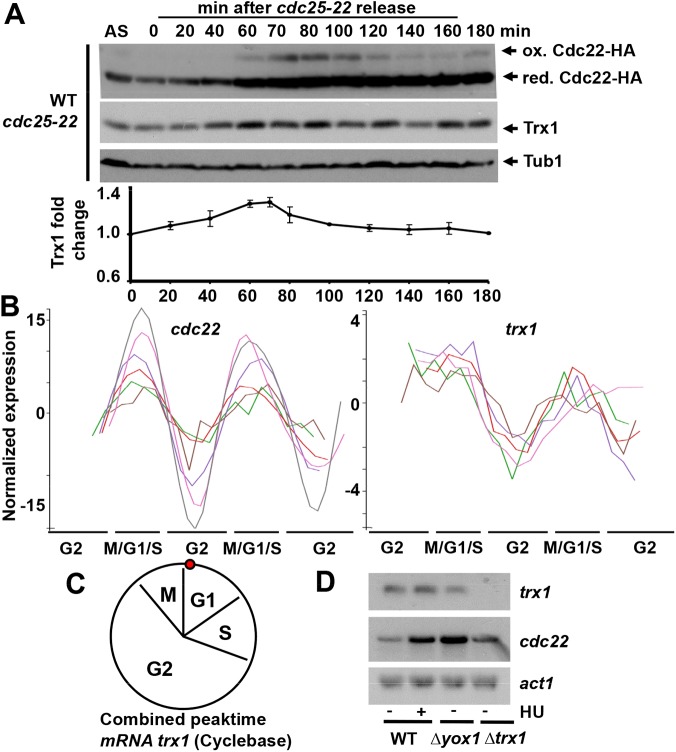
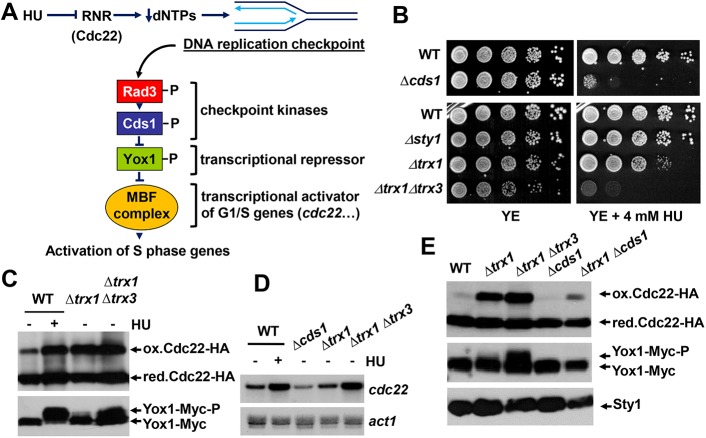
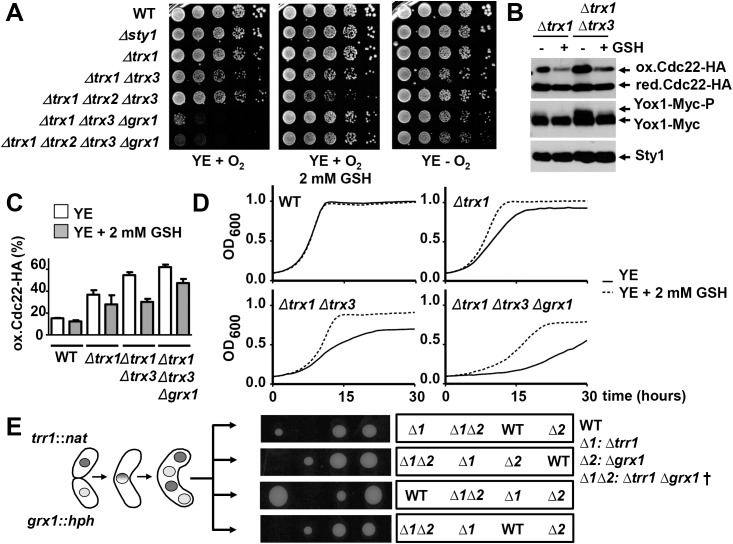
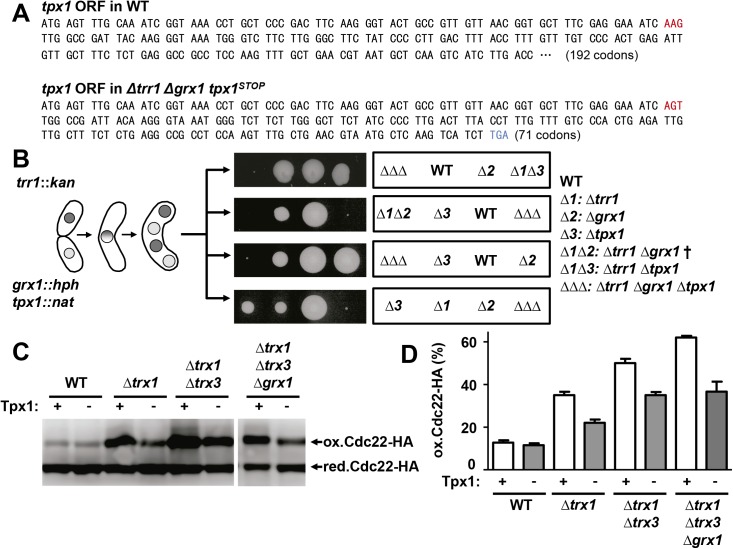
The thioredoxin and glutaredoxin pathways are responsible of recycling several enzymes which undergo intramolecular disulfide bond formation as part of their catalytic cycles such as the peroxide scavengers peroxiredoxins or the enzyme ribonucleotide reductase (RNR). RNR, the rate-limiting enzyme of deoxyribonucleotide synthesis, is an essential enzyme relying on these electron flow cascades for recycling. RNR is tightly regulated in a cell cycle-dependent manner at different levels, but little is known about the participation of electron donors in such regulation. Here, we show that cytosolic thioredoxins Trx1 and Trx3 are the primary electron donors for RNR in fission yeast. Unexpectedly, trx1 transcript and Trx1 protein levels are up-regulated in a G1-to-S phase-dependent manner, indicating that the supply of electron donors is also cell cycle-regulated. Indeed, genetic depletion of thioredoxins triggers a DNA replication checkpoint ruled by Rad3 and Cds1, with the final goal of up-regulating transcription of S phase genes and constitutive RNR synthesis. Regarding the thioredoxin and glutaredoxin cascades, one combination of gene deletions is synthetic lethal in fission yeast: cells lacking both thioredoxin reductase and cytosolic dithiol glutaredoxin. We have isolated a suppressor of this lethal phenotype: a mutation at the Tpx1-coding gene, leading to a frame shift and a loss-of-function of Tpx1, the main client of electron donors. We propose that in a mutant strain compromised in reducing equivalents, the absence of an abundant and competitive substrate such as the peroxiredoxin Tpx1 has been selected as a lethality suppressor to favor RNR function at the expense of the non-essential peroxide scavenging function, to allow DNA synthesis and cell growth.
硫氧还蛋白和谷氧还蛋白途径负责循环利用几种酶,这些酶在其催化循环中会形成分子内二硫键,例如过氧化物清除剂过氧化物酶或核糖核苷酸还原酶(RNR)。RNR是脱氧核糖核苷酸合成的限速酶,是一种依赖于这些电子流级联反应进行循环利用的必需酶。RNR在细胞周期的不同水平上受到严格的细胞周期依赖性调控,但关于电子供体在这种调控中的参与情况知之甚少。在这里,我们表明胞质硫氧还蛋白Trx1和Trx3是裂殖酵母中RNR的主要电子供体。出乎意料的是,trx1转录本和Trx1蛋白水平以G1到S期依赖性方式上调,表明电子供体的供应也受到细胞周期调控。实际上,硫氧还蛋白的基因缺失会触发由Rad3和Cds1控制的DNA复制检查点,最终目标是上调S期基因的转录和组成型RNR合成。关于硫氧还蛋白和谷氧还蛋白级联反应,一种基因缺失组合在裂殖酵母中是合成致死的:同时缺乏硫氧还蛋白还原酶和胞质二硫醇谷氧还蛋白的细胞。我们分离出了这种致死表型的一个抑制子:Tpx1编码基因的一个突变,导致移码和Tpx1(电子供体的主要底物)功能丧失。我们提出,在还原当量受损的突变菌株中,选择缺失一种丰富且有竞争力的底物(如过氧化物酶Tpx1)作为致死性抑制子,以牺牲非必需的过氧化物清除功能为代价来促进RNR功能,从而允许DNA合成和细胞生长。