Razin Sergey V, Ulianov Sergey V
Institute of Gene Biology, Russian Academy of Sciences, Vavilov Street 34/5, 119334 Moscow, Russia.
Lomonosov Moscow State University, Biological Faculty, Leninskie Gory 1, building 12, 119192 Moscow, Russia.
Cell Mol Biol Lett. 2017 Aug 29;22:18. doi: 10.1186/s11658-017-0050-4. eCollection 2017.
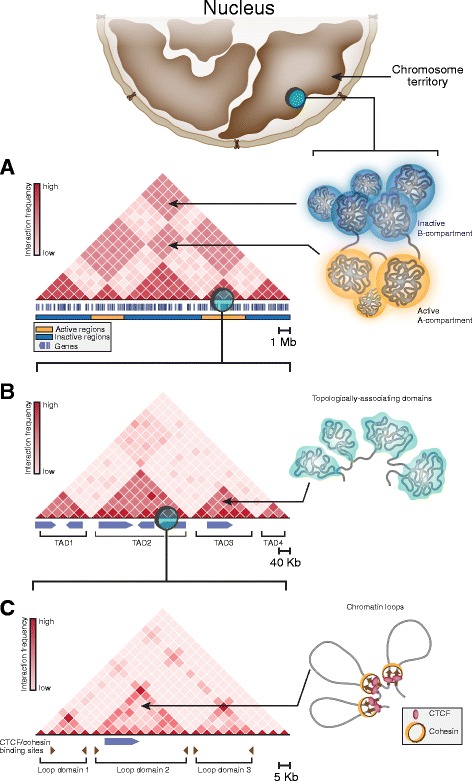
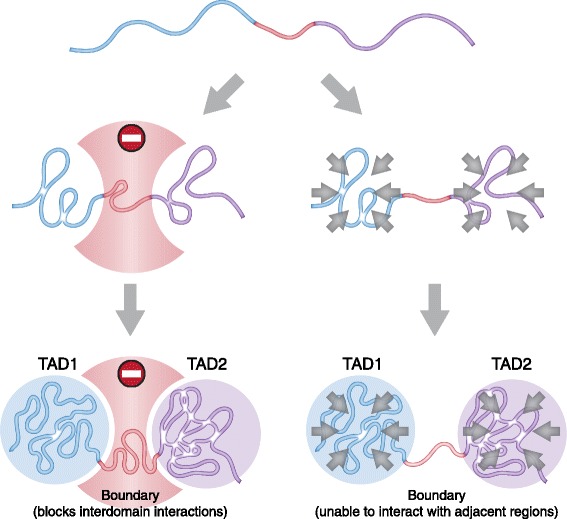
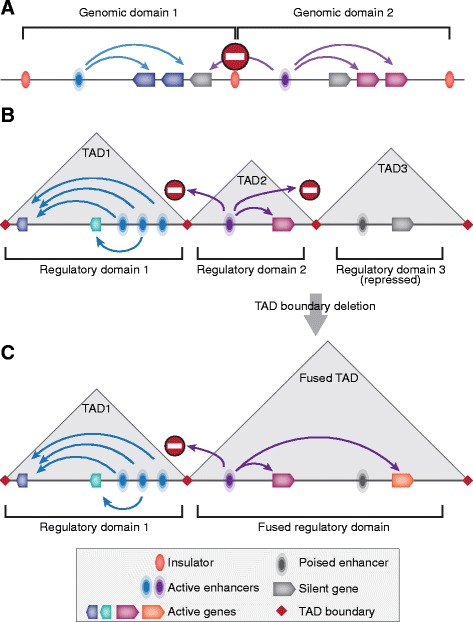
In mammals, genomic DNA that is roughly 2 m long is folded to fit the size of the cell nucleus that has a diameter of about 10 μm. The folding of genomic DNA is mediated via assembly of DNA-protein complex, chromatin. In addition to the reduction of genomic DNA linear dimensions, the assembly of chromatin allows to discriminate and to mark active (transcribed) and repressed (non-transcribed) genes. Consequently, epigenetic regulation of gene expression occurs at the level of DNA packaging in chromatin. Taking into account the increasing attention of scientific community toward epigenetic systems of gene regulation, it is very important to understand how DNA folding in chromatin is related to gene activity. For many years the hierarchical model of DNA folding was the most popular. It was assumed that nucleosome fiber (10-nm fiber) is folded into 30-nm fiber and further on into chromatin loops attached to a nuclear/chromosome scaffold. Recent studies have demonstrated that there is much less regularity in chromatin folding within the cell nucleus. The very existence of 30-nm chromatin fibers in living cells was questioned. On the other hand, it was found that chromosomes are partitioned into self-interacting spatial domains that restrict the area of enhancers action. Thus, TADs can be considered as structural-functional domains of the chromosomes. Here we discuss the modern view of DNA packaging within the cell nucleus in relation to the regulation of gene expression. Special attention is paid to the possible mechanisms of the chromatin fiber self-assembly into TADs. We discuss the model postulating that partitioning of the chromosome into TADs is determined by the distribution of active and inactive chromatin segments along the chromosome. This article was specially invited by the editors and represents work by leading researchers.
在哺乳动物中,大约2米长的基因组DNA被折叠起来,以适应直径约为10微米的细胞核的大小。基因组DNA的折叠是通过DNA-蛋白质复合物(染色质)的组装来介导的。除了减小基因组DNA的线性尺寸外,染色质的组装还能够区分并标记活跃(转录)和受抑制(非转录)的基因。因此,基因表达的表观遗传调控发生在染色质中DNA包装的水平上。考虑到科学界对基因调控的表观遗传系统越来越关注,了解染色质中的DNA折叠与基因活性之间的关系非常重要。多年来,DNA折叠的层次模型最为流行。人们认为核小体纤维(10纳米纤维)折叠成30纳米纤维,进而折叠成附着在核/染色体支架上的染色质环。最近的研究表明,细胞核内染色质折叠的规律性要低得多。活细胞中30纳米染色质纤维的存在受到质疑。另一方面,人们发现染色体被划分为自我相互作用的空间结构域,这些结构域限制了增强子的作用区域。因此,拓扑相关结构域(TADs)可被视为染色体的结构功能域。在这里,我们讨论关于细胞核内DNA包装与基因表达调控相关的现代观点。特别关注染色质纤维自组装成TADs的可能机制。我们讨论了一种模型,该模型假设染色体划分为TADs是由活跃和不活跃染色质片段沿染色体的分布决定的。本文是由编辑特别邀请撰写的,代表了顶尖研究人员的工作。