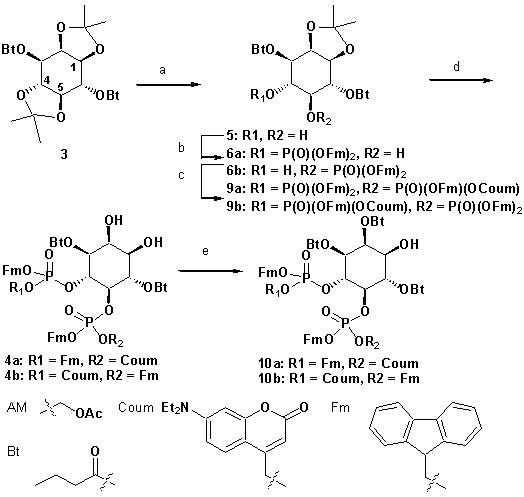
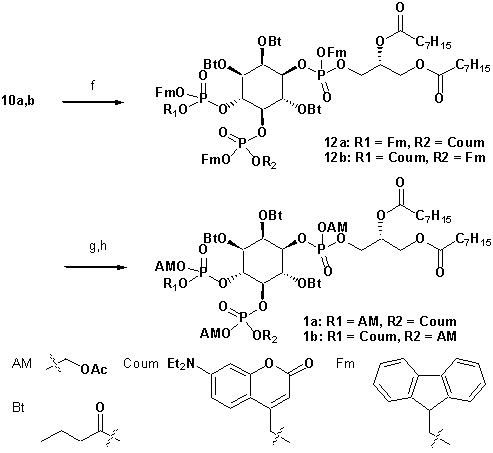
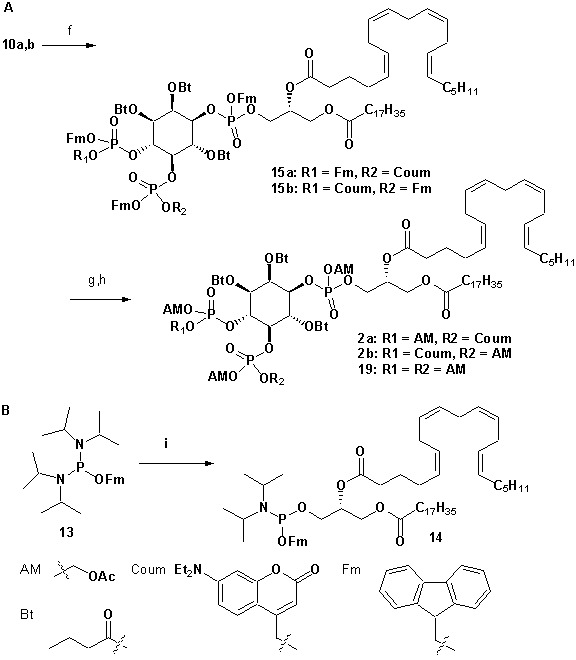
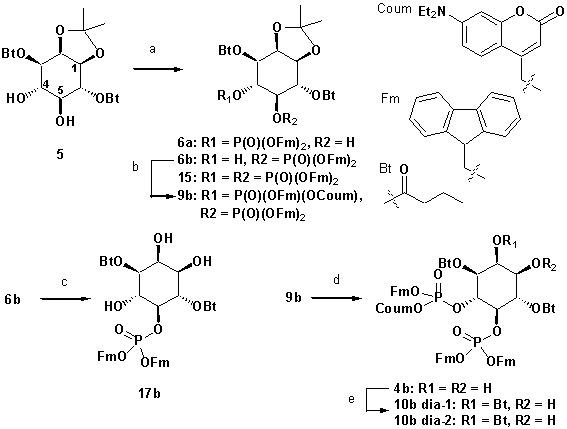
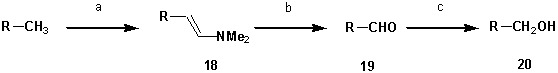Neurosecretion group, Center for Neuroscience, Faculty of Health and Medical Sciences, University of Copenhagen, Copenhagen, Denmark.
Leibniz-Forschungsinstitut für Molekulare Pharmakologie, Berlin, Germany.
Elife. 2017 Oct 25;6:e30203. doi: 10.7554/eLife.30203.
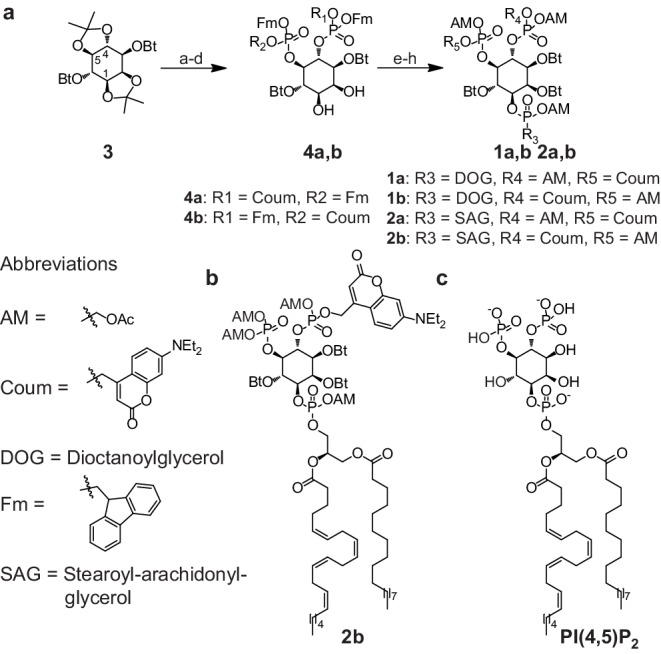
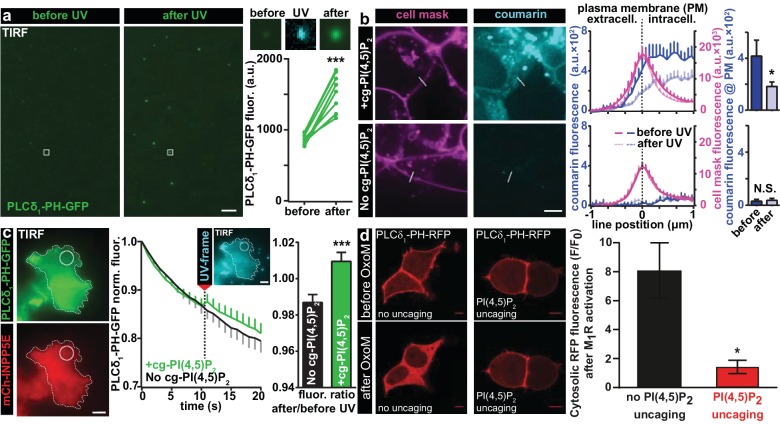
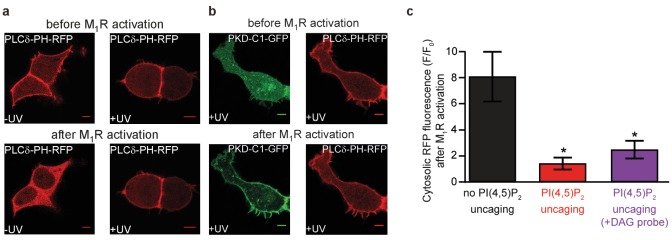
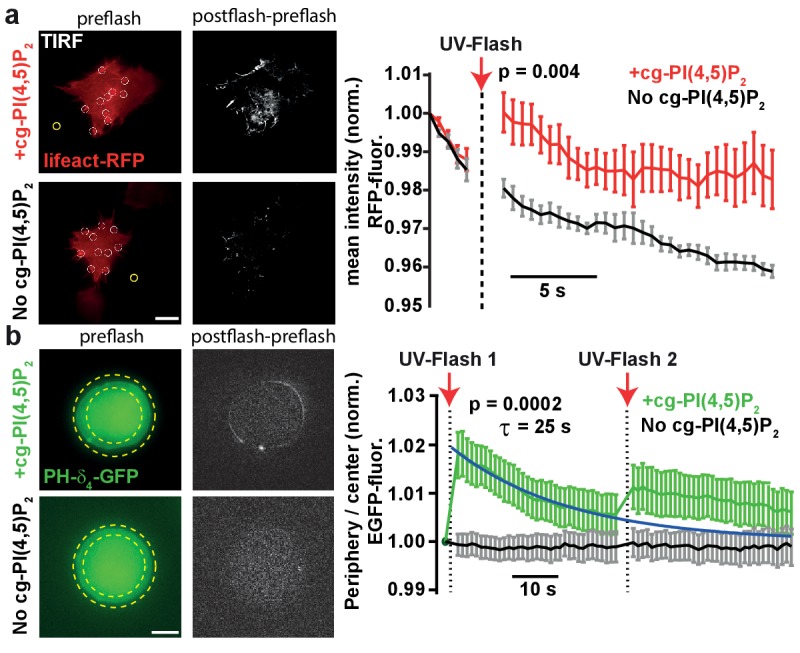
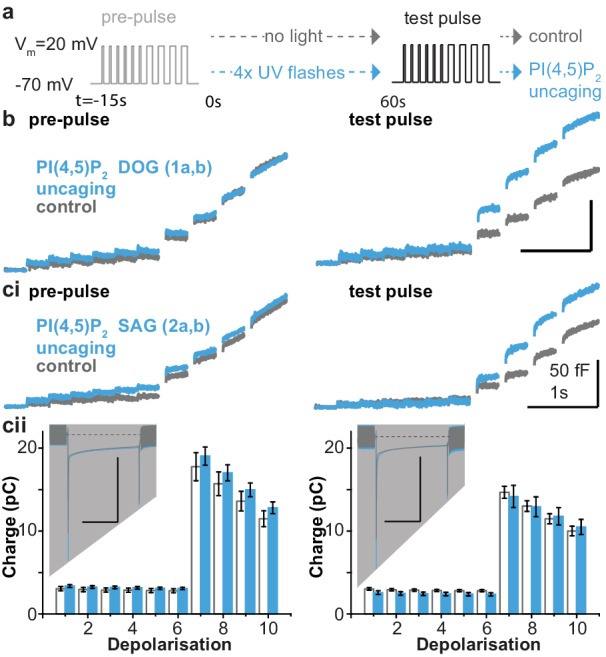
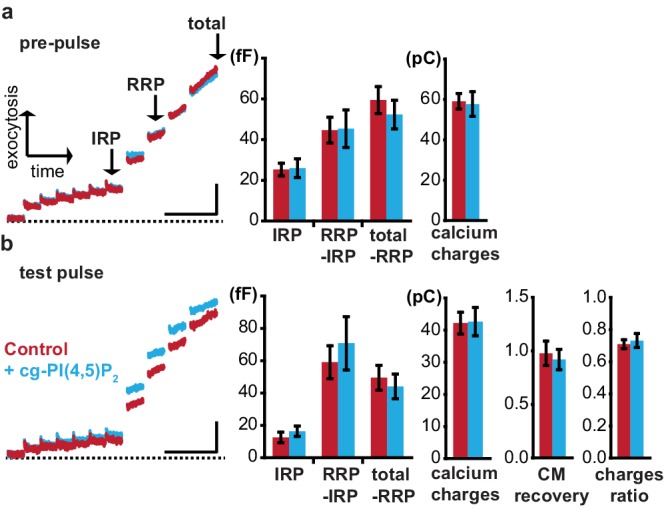
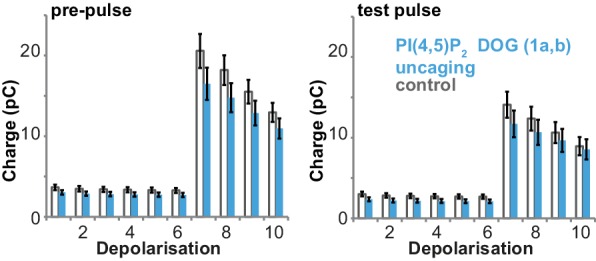
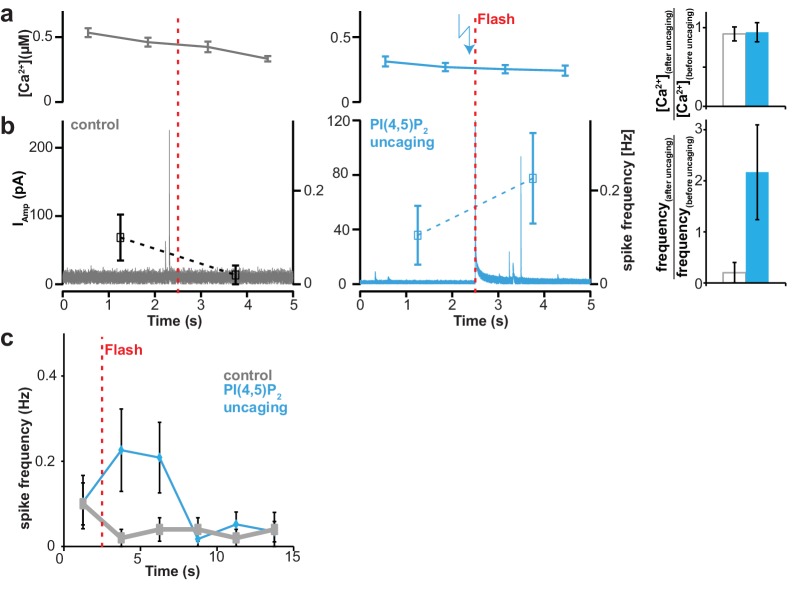
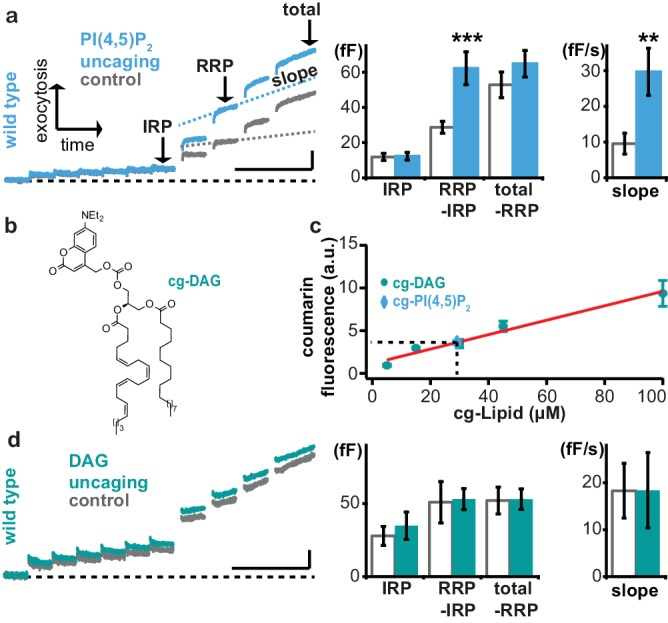
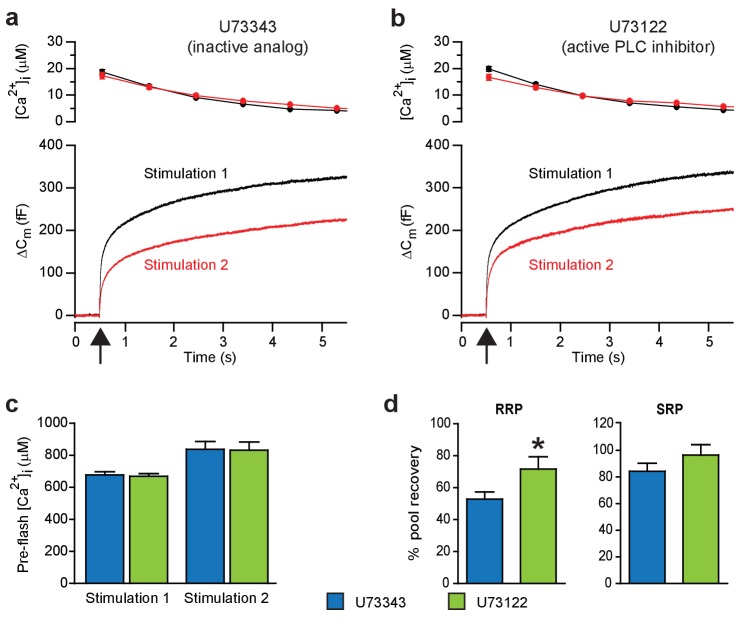
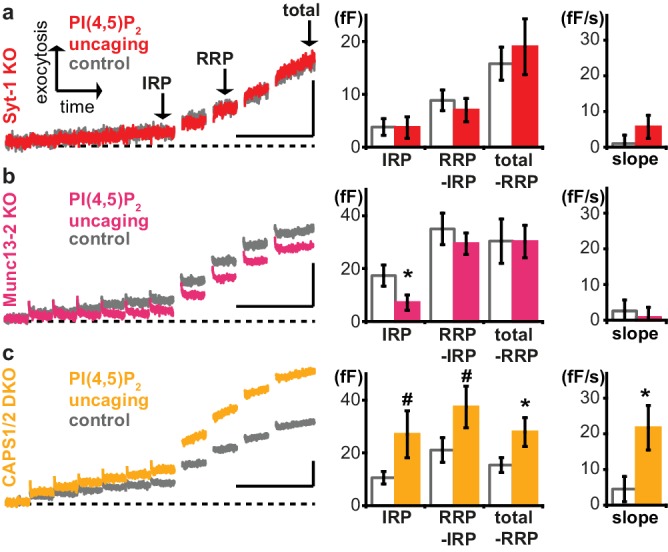
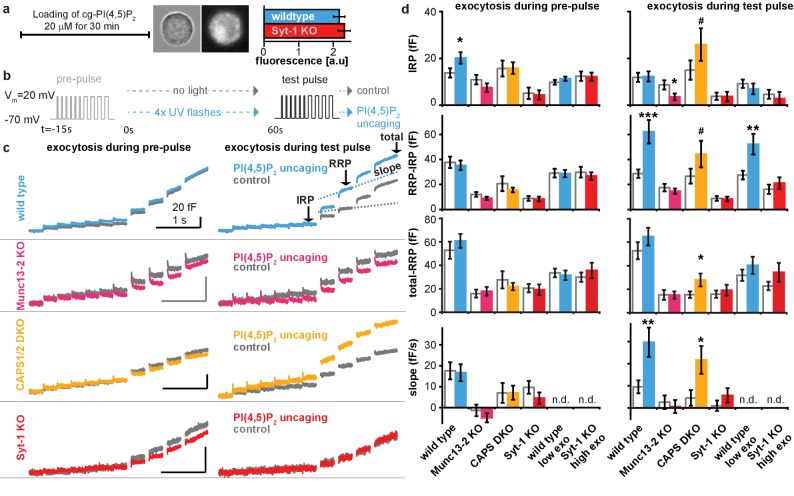
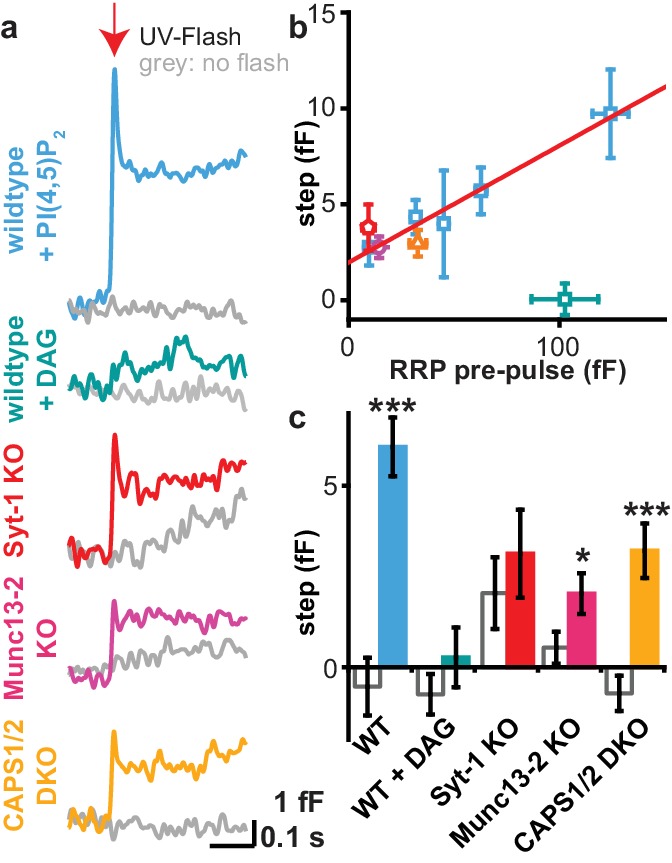
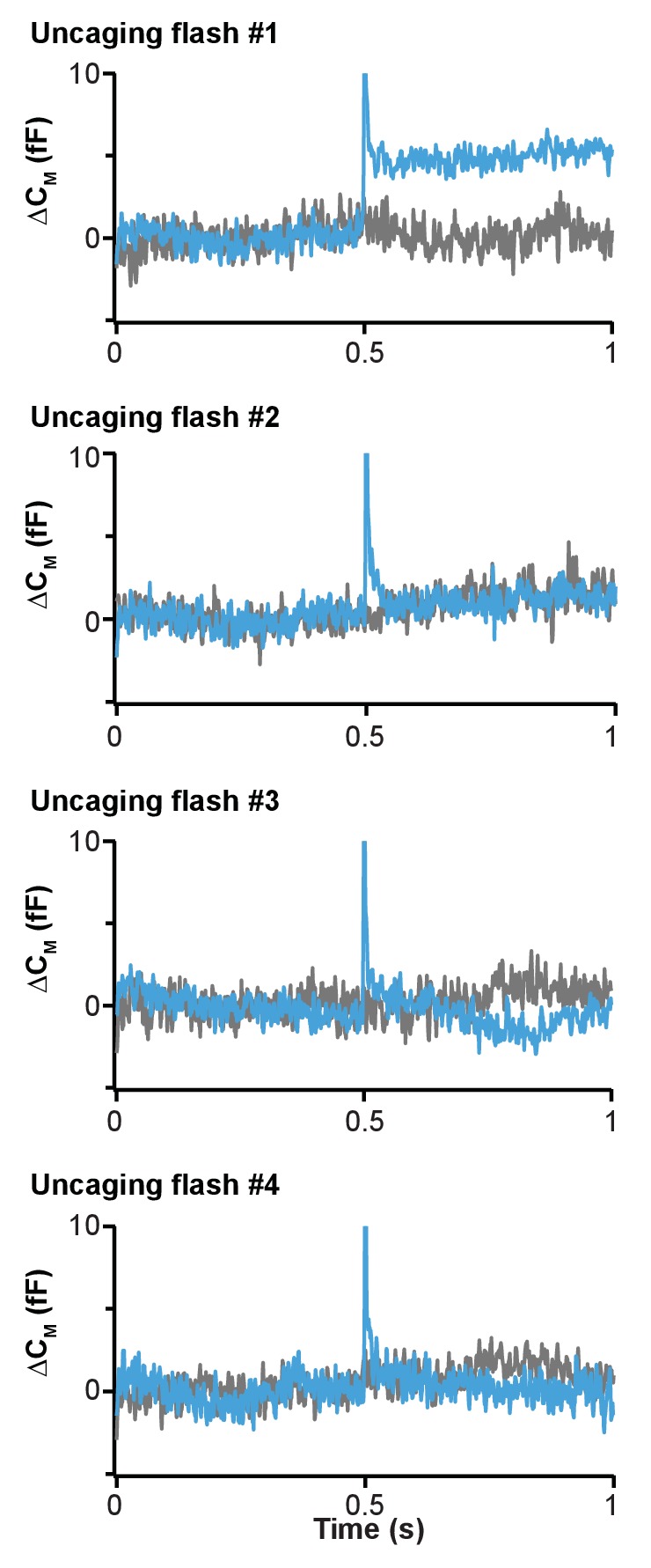
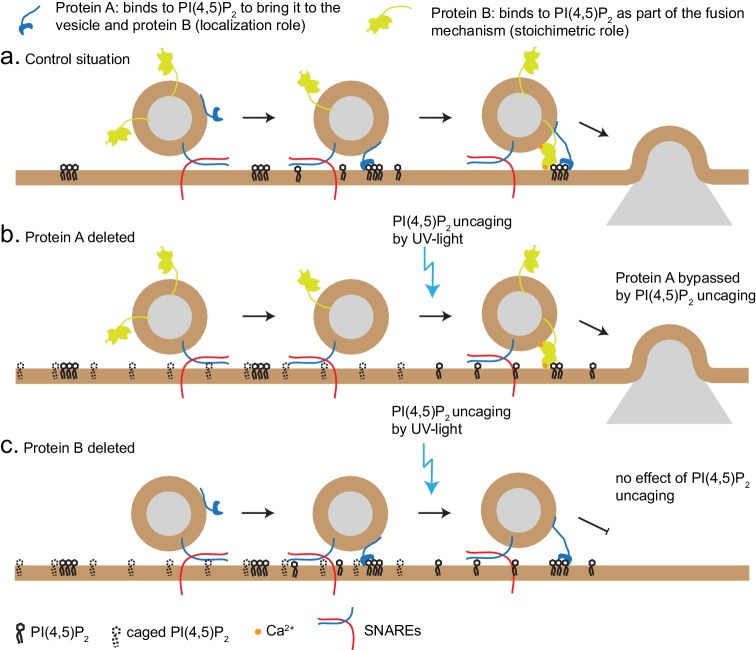
Phosphatidylinositol-4,5-bisphosphate [PI(4,5)P] is essential for exocytosis. Classical ways of manipulating PI(4,5)P levels are slower than its metabolism, making it difficult to distinguish effects of PI(4,5)P from those of its metabolites. We developed a membrane-permeant, photoactivatable PI(4,5)P, which is loaded into cells in an inactive form and activated by light, allowing sub-second increases in PI(4,5)P levels. By combining this compound with electrophysiological measurements in mouse adrenal chromaffin cells, we show that PI(4,5)P uncaging potentiates exocytosis and identify synaptotagmin-1 (the Ca sensor for exocytosis) and Munc13-2 (a vesicle priming protein) as the relevant effector proteins. PI(4,5)P activation of exocytosis did not depend on the PI(4,5)P-binding CAPS-proteins, suggesting that PI(4,5)P uncaging may bypass CAPS-function. Finally, PI(4,5)P uncaging triggered the rapid fusion of a subset of readily-releasable vesicles, revealing a rapid role of PI(4,5)P in fusion triggering. Thus, optical uncaging of signaling lipids can uncover their rapid effects on cellular processes and identify lipid effectors.
磷脂酰肌醇-4,5-二磷酸[PI(4,5)P]对于胞吐作用至关重要。经典的操纵 PI(4,5)P 水平的方法比其代谢速度慢,这使得难以区分 PI(4,5)P 的作用与其代谢物的作用。我们开发了一种膜通透的光活化 PI(4,5)P,它以非活性形式被加载到细胞中,并通过光激活,允许 PI(4,5)P 水平在亚秒内增加。通过将这种化合物与小鼠肾上腺嗜铬细胞中的电生理测量相结合,我们表明 PI(4,5)P 光解可增强胞吐作用,并确定突触结合蛋白-1(胞吐作用的 Ca 传感器)和 Munc13-2(囊泡引发蛋白)是相关的效应蛋白。PI(4,5)P 激活胞吐作用不依赖于 PI(4,5)P 结合 CAPS 蛋白,这表明 PI(4,5)P 光解可能绕过 CAPS 功能。最后,PI(4,5)P 光解触发了一组易于释放的囊泡的快速融合,揭示了 PI(4,5)P 在融合触发中的快速作用。因此,信号脂质的光解可以揭示它们对细胞过程的快速影响,并确定脂质效应物。