a Institute for Computational Medicine, Department of Biomedical Engineering, Institute for NanoBioTechnology , Johns Hopkins University , Baltimore , MD , USA.
Cell Adh Migr. 2018 May 4;12(3):204-214. doi: 10.1080/19336918.2017.1372878. Epub 2017 Nov 2.
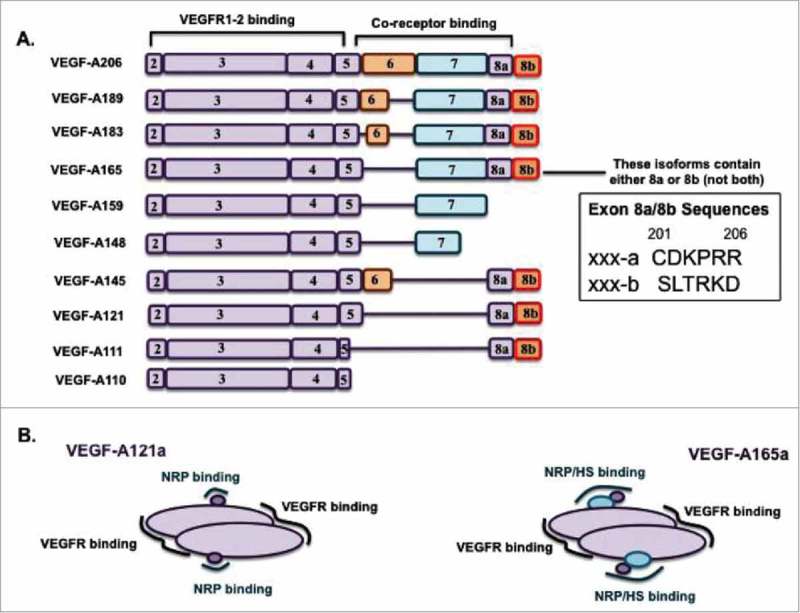
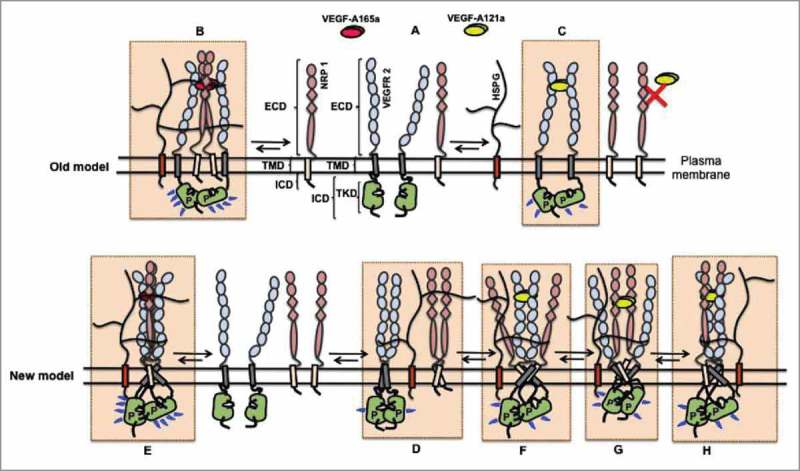
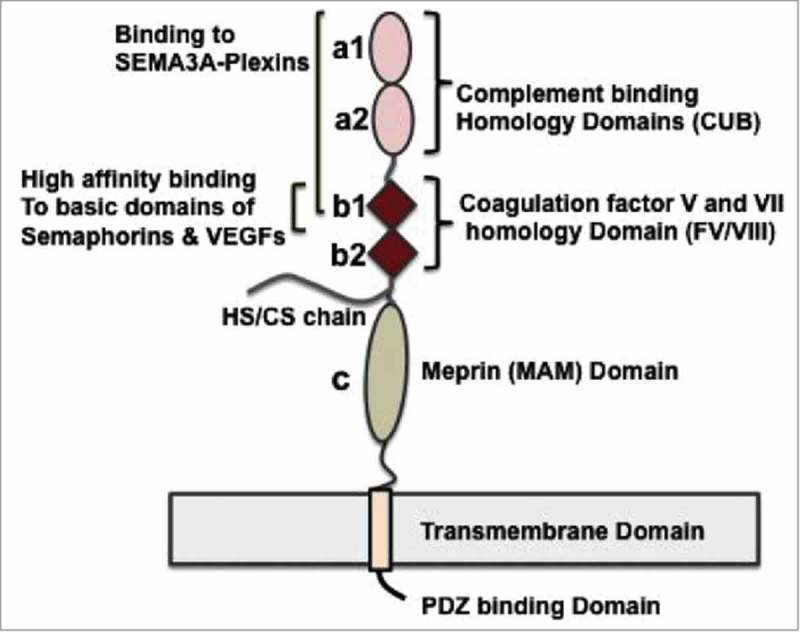
All known splice isoforms of vascular endothelial growth factor A (VEGF-A) can bind to the receptor tyrosine kinases VEGFR-1 and VEGFR-2. We focus here on VEGF-A121a and VEGF-A165a, two of the most abundant VEGF-A splice isoforms in human tissue , and their ability to bind the Neuropilin co-receptors NRP1 and NRP2. The Neuropilins are key vascular, immune, and nervous system receptors on endothelial cells, neuronal axons, and regulatory T cells respectively. They serve as co-receptors for the Plexins in Semaphorin binding on neuronal and vascular endothelial cells, and for the VEGFRs in VEGF binding on vascular and lymphatic endothelial cells, and thus regulate the initiation and coordination of cell signaling by Semaphorins and VEGFs. There is conflicting evidence in the literature as to whether only heparin-binding VEGF-A isoforms - that is, isoforms with domains encoded by exons 6 and/or 7 plus 8a - bind to Neuropilins on endothelial cells. While it is clear that VEGF-A165a binds to both NRP1 and NRP2, published studies do not all agree on the ability of VEGF-A121a to bind NRPs. Here, we review and attempt to reconcile evidence for and against VEGF-A121a binding to Neuropilins. This evidence suggests that, in vitro, VEGF-A121a can bind to both NRP1 and NRP2 via domains encoded by exons 5 and 8a; in the case of NRP1, VEGF-A121a binds with lower affinity than VEGF-A165a. In in vitro cell culture experiments, both NRP1 and NRP2 can enhance VEGF-A121a-induced phosphorylation of VEGFR2 and downstream signaling including proliferation. However, unlike VEGFA-165a, experiments have shown that VEGF-A121a does not 'bridge' VEGFR2 and NRP1, i.e. it does not bind both receptors simultaneously at their extracellular domain. Thus, the mechanism by which Neuropilins potentiate VEGF-A121a-mediated VEGFR2 signaling may be different from that for VEGF-A165a. We suggest such an alternate mechanism: interactions between NRP1 and VEGFR2 transmembrane (TM) and intracellular (IC) domains.
所有已知的血管内皮生长因子 A(VEGF-A)剪接异构体都可以与受体酪氨酸激酶 VEGFR-1 和 VEGFR-2 结合。我们在这里关注的是 VEGF-A121a 和 VEGF-A165a,这两种是人类组织中最丰富的 VEGF-A 剪接异构体之一,以及它们与神经纤毛蛋白共受体 NRP1 和 NRP2 结合的能力。神经纤毛蛋白分别是内皮细胞、神经元轴突和调节性 T 细胞的关键血管、免疫和神经系统受体。它们作为神经丝素在神经元和血管内皮细胞上结合的 Plexin 的共受体,以及 VEGF 在血管和淋巴管内皮细胞上结合的 VEGFR 的共受体,从而调节神经丝素和 VEGF 的细胞信号的启动和协调。文献中有相互矛盾的证据表明,只有肝素结合的 VEGF-A 异构体,即具有外显子 6 和/或 7 以及 8a 编码的结构域的异构体,才能与内皮细胞上的神经纤毛蛋白结合。虽然 VEGF-A165a 显然与 NRP1 和 NRP2 都结合,但已发表的研究并不都同意 VEGF-A121a 与 NRPs 结合的能力。在这里,我们回顾并试图调和支持和反对 VEGF-A121a 与神经纤毛蛋白结合的证据。这些证据表明,在体外,VEGF-A121a 可以通过外显子 5 和 8a 编码的结构域与 NRP1 和 NRP2 结合;在 NRP1 的情况下,VEGF-A121a 的结合亲和力低于 VEGF-A165a。在体外细胞培养实验中,NRP1 和 NRP2 都可以增强 VEGF-A121a 诱导的 VEGFR2 磷酸化和下游信号转导,包括增殖。然而,与 VEGFA-165a 不同的是,实验表明 VEGF-A121a 不能“桥接”VEGFR2 和 NRP1,即它不能同时在其细胞外结构域结合两个受体。因此,神经纤毛蛋白增强 VEGF-A121a 介导的 VEGFR2 信号转导的机制可能与 VEGF-A165a 不同。我们提出了一种替代机制:NRP1 和 VEGFR2 跨膜(TM)和细胞内(IC)结构域之间的相互作用。