Department of Microbiology and Immunology, Weill Cornell Medical College, New York, New York 10065, USA.
Molecular Biology Program, Sloan-Kettering Institute, New York, New York 10065, USA.
RNA. 2018 Feb;24(2):237-250. doi: 10.1261/rna.063966.117. Epub 2017 Nov 9.
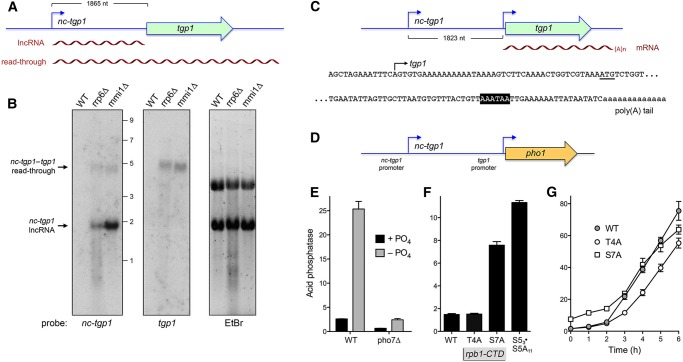
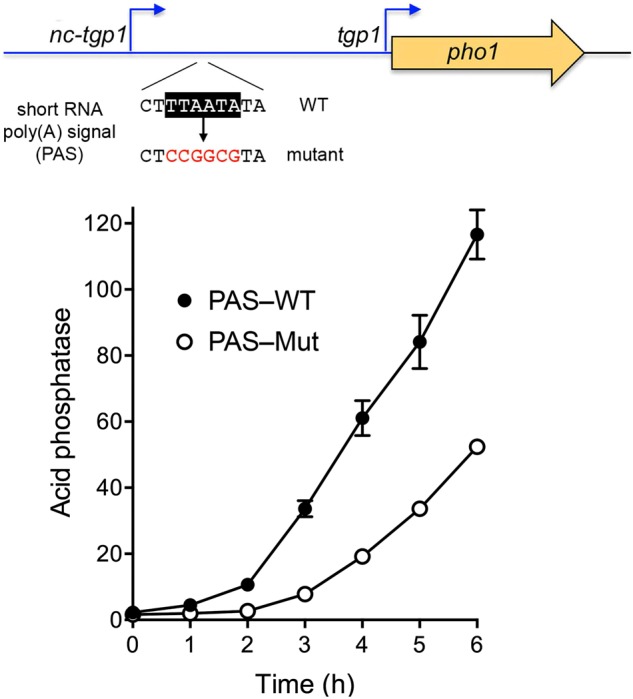
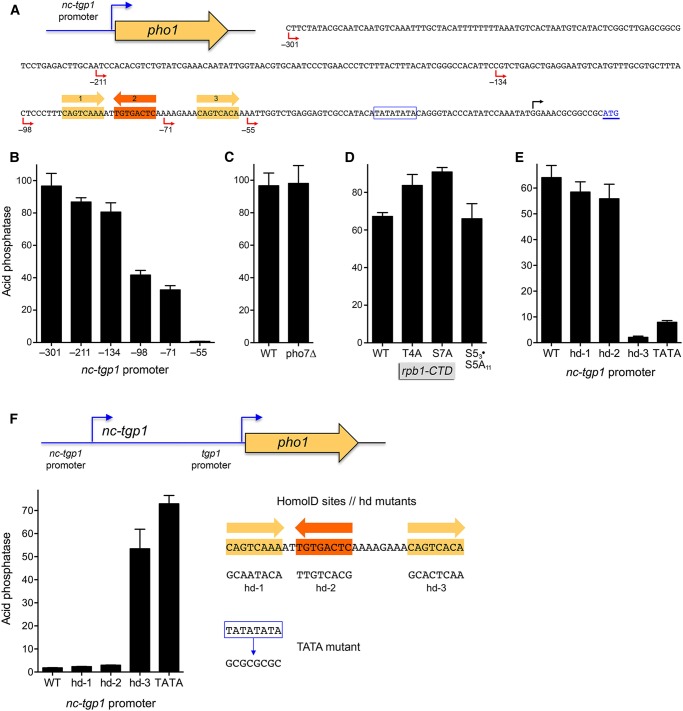
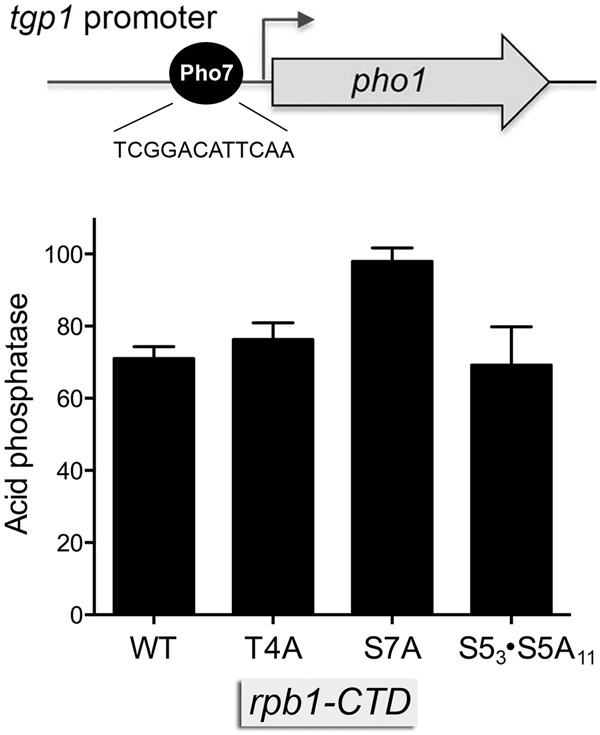
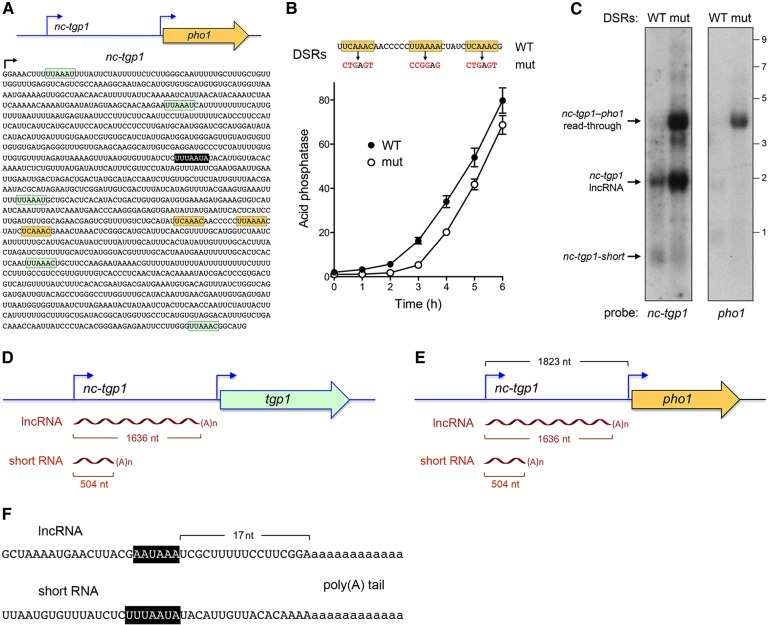
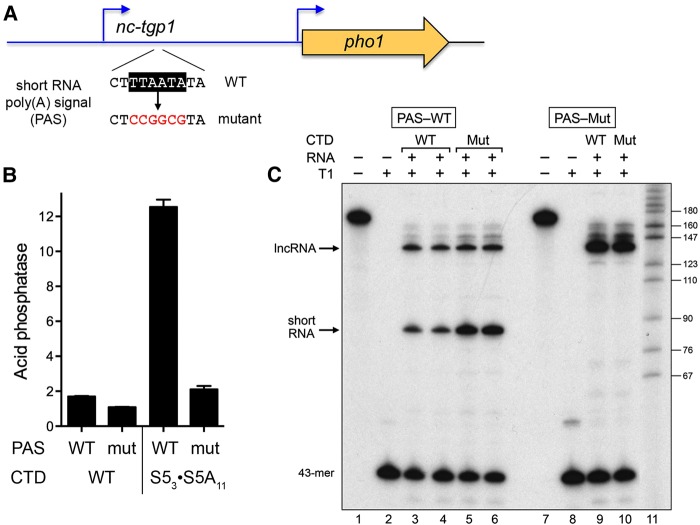
Expression of fission yeast glycerophosphate transporter Tgp1 is repressed in phosphate-rich medium and induced during phosphate starvation. Repression is enforced by transcription of the locus upstream of to produce a long noncoding (lnc) RNA. Here we identify two essential elements of the promoter: a TATA box TATATATA and a HomolD box CAGTCACA, mutations of which inactivate the promoter and de-repress the downstream promoter under phosphate-replete conditions. The lncRNA poly(A) site maps to nucleotide +1636 of the transcription unit, which coincides with the binding site for Pho7 (TCGGACATTCAA), the transcription factor that drives expression. Overlap between the lncRNA template and the promoter points to transcriptional interference as the simplest basis for lncRNA repression. We identify a shorter RNA derived from the locus, polyadenylated at position +508, well upstream of the promoter. Mutating the RNA polyadenylation signal abolishes de-repression of the downstream promoter elicited by Pol2 CTD Ser5Ala phospho-site mutation. Ser5 mutation favors utilization of the short RNA poly(A) site, thereby diminishing transcription of the lncRNA that interferes with the promoter. Mutating the RNA polyadenylation signal attenuates induction of the promoter during phosphate starvation. Polyadenylation site choice governed by CTD Ser5 status adds a new level of lncRNA control of gene expression and reveals a new feature of the fission yeast CTD code.
裂殖酵母甘油磷酸转运蛋白 Tgp1 的表达在富含磷酸盐的培养基中受到抑制,在磷酸盐饥饿时被诱导。这种抑制是通过转录位于 Tgp1 上游的 基因座产生长非编码 (lnc) RNA 来实现的。在这里,我们确定了 启动子的两个必需元件:一个 TATA 盒 TATATATA 和一个 HomolD 盒 CAGTCACA,它们的突变使 启动子失活,并在磷酸盐充足的条件下解除对下游 启动子的抑制。lncRNA 的 poly(A) 位点映射到转录单位的+1636 核苷酸,与驱动 表达的转录因子 Pho7(TCGGACATTCAA)的结合位点相吻合。lncRNA 模板与 启动子之间的重叠表明转录干扰是 lncRNA 抑制的最简单基础。我们鉴定了来自 基因座的较短 RNA,其 poly(A) 位点位于+508 位,远在 启动子的上游。突变 RNA 的 poly(A) 信号会破坏 Pol2 CTD Ser5Ala 磷酸化位点突变引起的下游 启动子的去抑制。Ser5 突变有利于利用较短的 RNA poly(A) 位点,从而减少干扰 启动子转录的 lncRNA 的转录。突变 RNA 的 poly(A) 信号减弱了在磷酸盐饥饿时 启动子的诱导。CTD Ser5 状态控制的多聚腺苷酸化位点选择为基因表达的 lncRNA 调控增加了一个新的层次,并揭示了裂殖酵母 CTD 密码的一个新特征。