Molecular Biology Program, Memorial Sloan Kettering Cancer Centergrid.51462.34, New York, New York, USA.
Gerstner Sloan Kettering Graduate School of Biomedical Sciences, New York, New York, USA.
mBio. 2022 Jun 28;13(3):e0103422. doi: 10.1128/mbio.01034-22. Epub 2022 May 10.
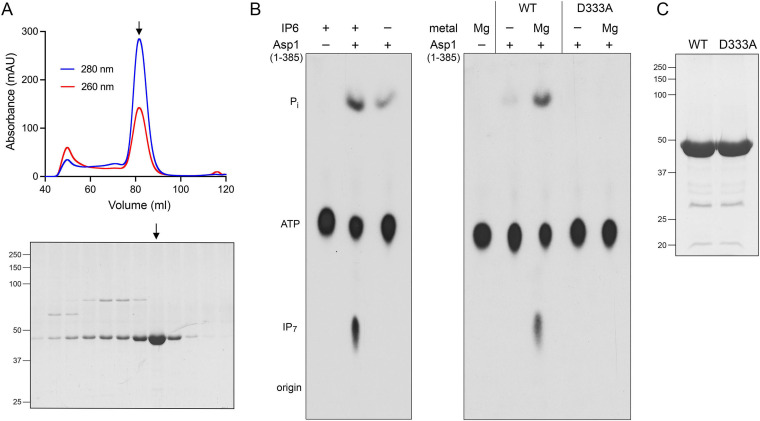
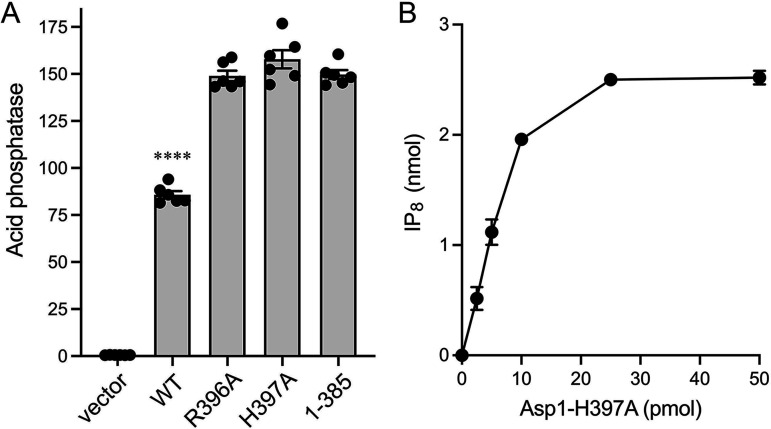
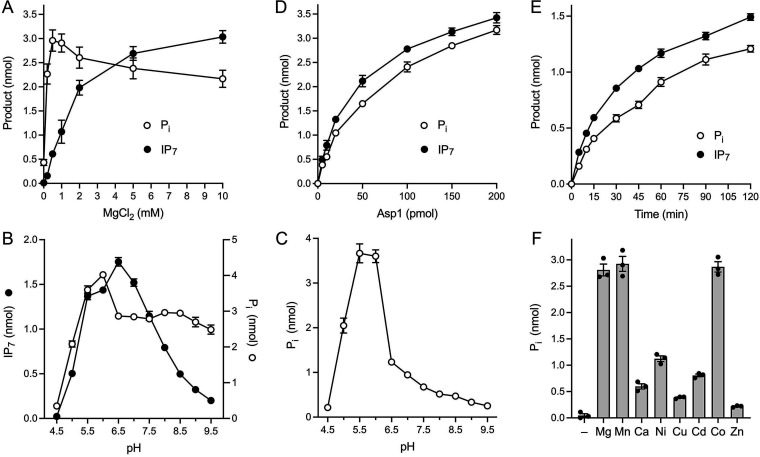
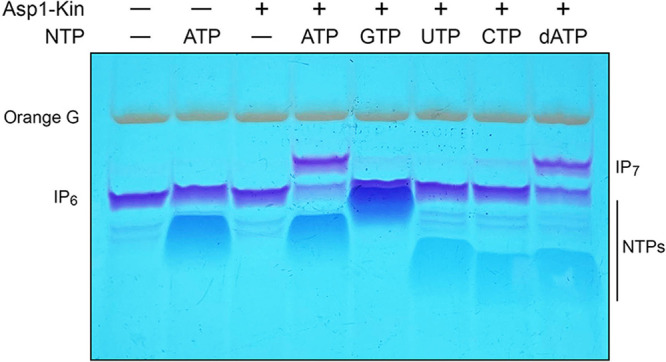
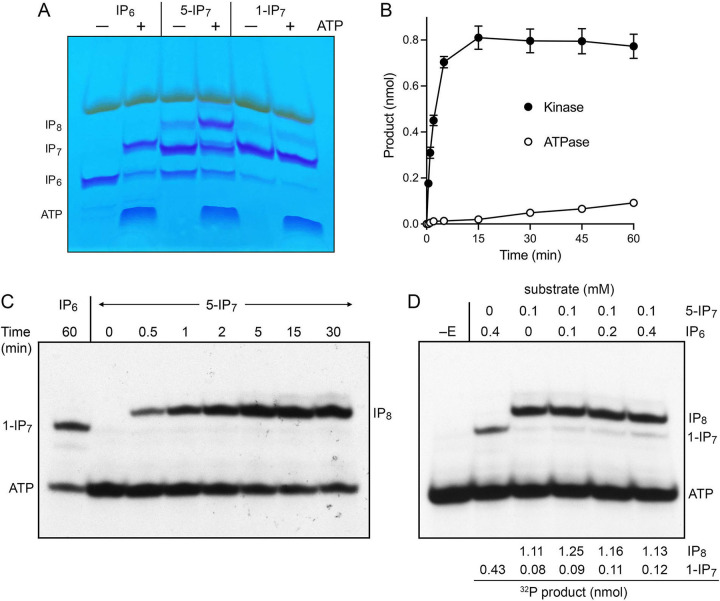
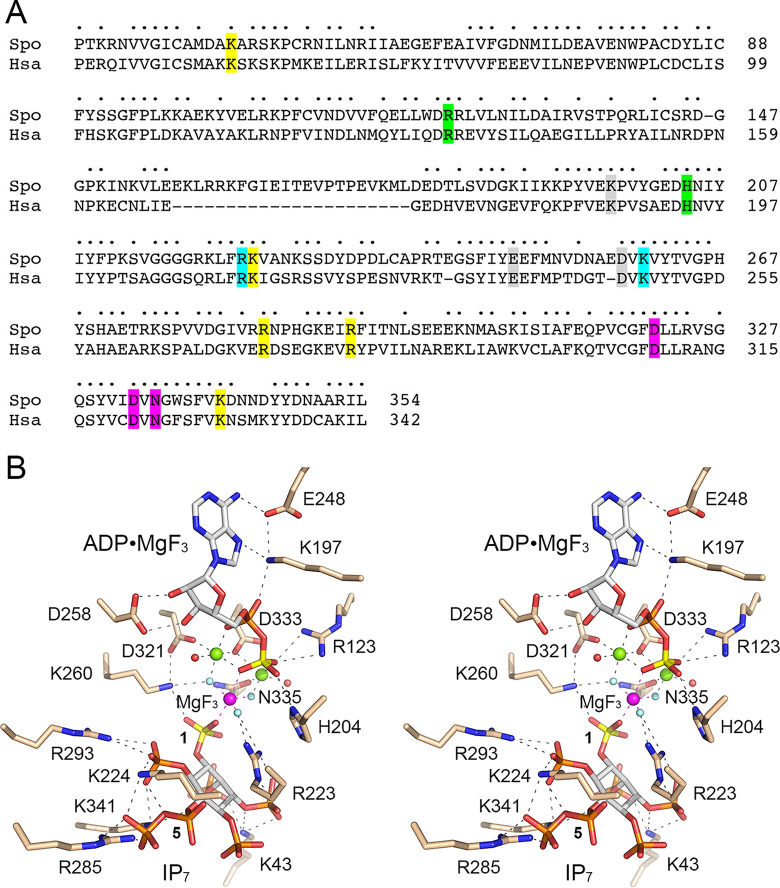
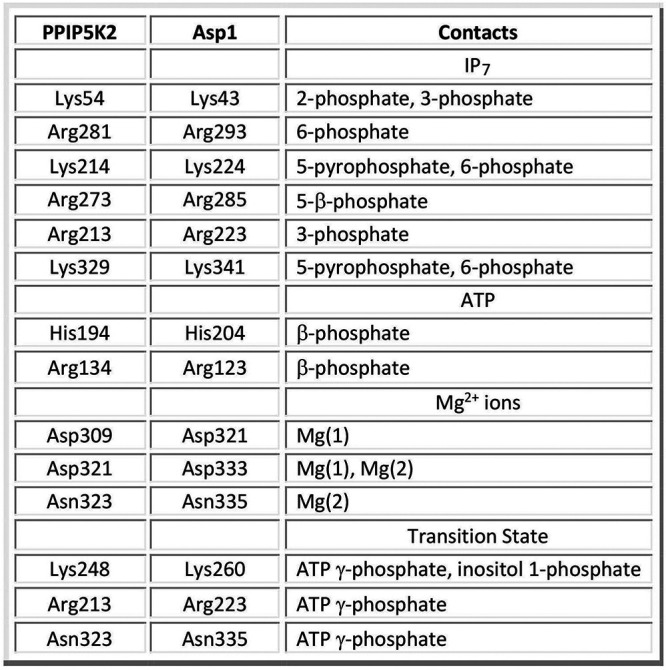
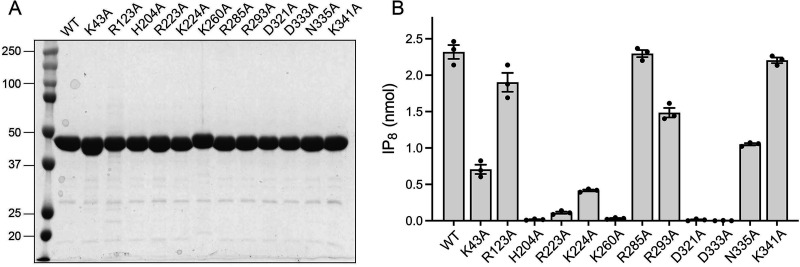
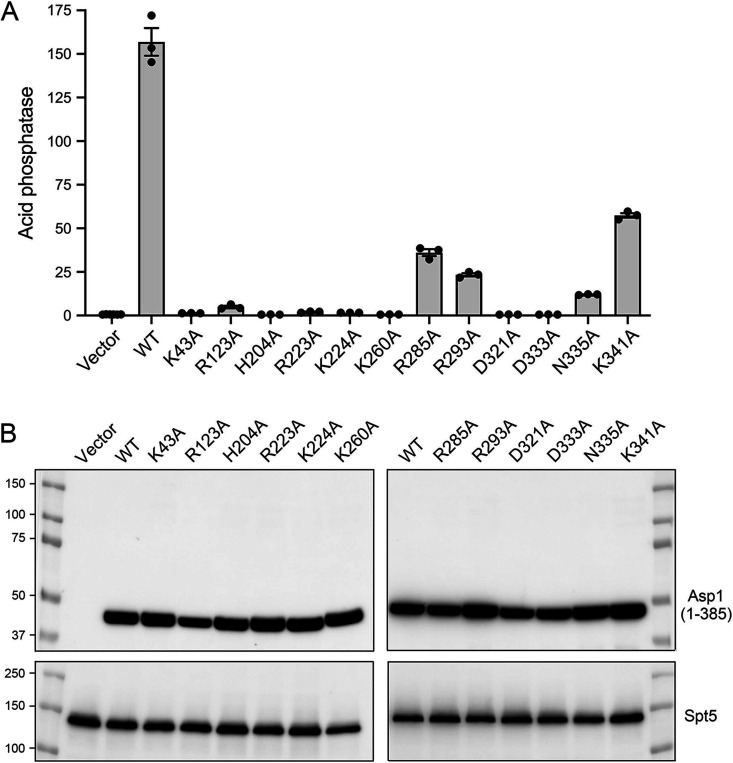
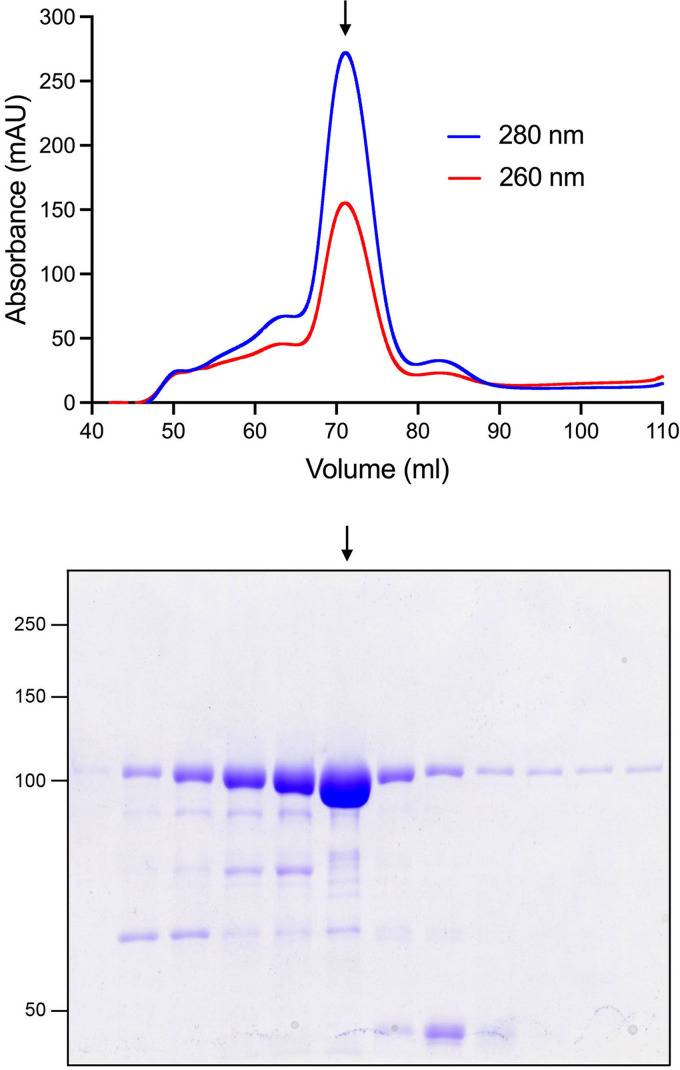
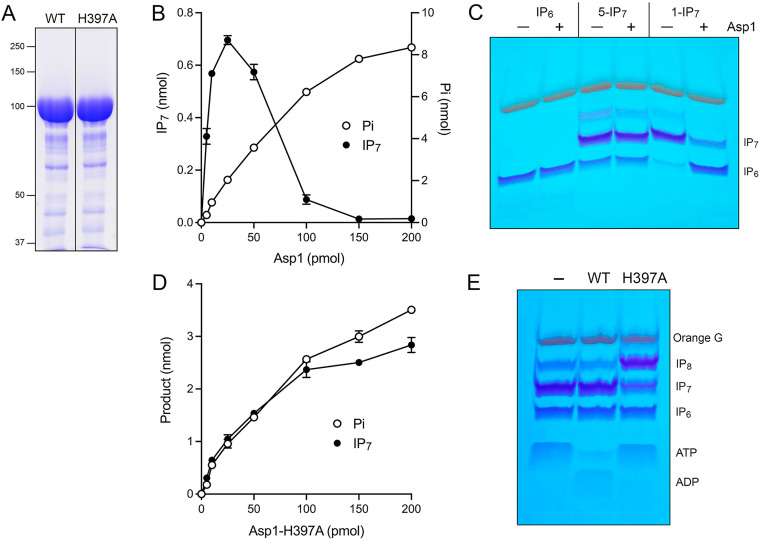
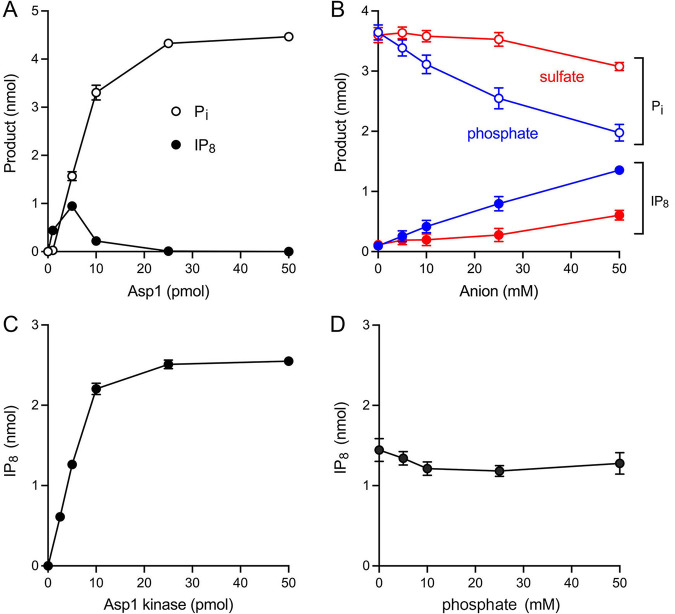
Inositol pyrophosphates (IPPs) are signaling molecules that regulate cellular phosphate homeostasis in diverse eukaryal taxa. In fission yeast, mutations that increase 1,5-IP derepress the regulon while mutations that ablate IP synthesis are hyper-repressive. Fission yeast Asp1, the principal agent of 1,5-IP dynamics, is a bifunctional enzyme composed of an N-terminal IPP kinase domain and a C-terminal IPP pyrophosphatase domain. Here we conducted a biochemical characterization and mutational analysis of the autonomous Asp1 kinase domain (aa 1-385). Reaction of Asp1 kinase with IP and ATP resulted in both IP phosphorylation to 1-IP and hydrolysis of the ATP γ-phosphate, with near-equal partitioning between productive 1-IP synthesis and unproductive ATP hydrolysis under optimal kinase conditions. By contrast, reaction of Asp1 kinase with 5-IP is 22-fold faster than with IP and is strongly biased in favor of IP synthesis versus ATP hydrolysis. Alanine scanning identified essential constituents of the active site. We deployed the Ala mutants to show that derepression of expression correlated with Asp1's kinase activity. In the case of full-length Asp1, the activity of the C-terminal pyrophosphatase domain stifled net phosphorylation of the 1-position during reaction of Asp1 with ATP and either IP or 5-IP. We report that inorganic phosphate is a concentration-dependent enabler of net IP synthesis by full-length Asp1 , by virtue of its antagonism of IP turnover. Expression of the fission yeast phosphate regulon is sensitive to the intracellular level of the inositol pyrophosphate (IPP) signaling molecule 1,5-IP. IP dynamics are determined by Asp1, a bifunctional enzyme comprising N-terminal IPP 1-kinase and C-terminal IPP 1-pyrophosphatase domains that catalyze IP synthesis and catabolism, respectively. Here, we interrogated the activities and specificities of the Asp1 kinase domain and full length Asp1. We find that reaction of Asp1 kinase with 5-IP is 22-fold faster than with IP and is strongly biased in favor of IP synthesis versus the significant unproductive ATP hydrolysis seen during its reaction with IP. We report that full-length Asp1 catalyzes futile cycles of 1-phosphate phosphorylation by its kinase component and 1-pyrophosphate hydrolysis by its pyrophosphatase component that result in unproductive net consumption of the ATP substrate. Net synthesis of 1,5-IP is enabled by physiological concentrations of inorganic phosphate that selectively antagonize IP turnover.
肌醇六磷酸(IPPs)是调节各种真核生物细胞内磷酸盐稳态的信号分子。在裂殖酵母中,增加 1,5-IPP 的突变会使 调控子去抑制,而消除 IPP 合成的突变则是高度抑制的。裂殖酵母的 Asp1 是 1,5-IPP 动力学的主要调节剂,它是一种由 N 端 IPP 激酶结构域和 C 端 IPP 焦磷酸酶结构域组成的双功能酶。在这里,我们对自主的 Asp1 激酶结构域(aa 1-385)进行了生化表征和突变分析。Asp1 激酶与 IP 和 ATP 的反应导致 IP 磷酸化生成 1-IP 和 ATP γ-磷酸水解,在最佳激酶条件下,产物 1-IP 合成和非产物 ATP 水解之间几乎相等。相比之下,Asp1 激酶与 5-IP 的反应速度比与 IP 的反应速度快 22 倍,并且强烈偏向于 IP 合成而不是 ATP 水解。丙氨酸扫描确定了活性位点的必需成分。我们利用 Ala 突变体表明, 的表达去抑制与 Asp1 的激酶活性相关。对于全长 Asp1,C 端焦磷酸酶结构域的活性在 Asp1 与 ATP 以及 IP 或 5-IP 反应过程中抑制了 1-位的净磷酸化。我们报告说,由于无机磷酸盐对 IP 转化的拮抗作用,它是全长 Asp1 净 IP 合成的浓度依赖性促进剂。裂殖酵母磷酸盐调控子的表达对肌醇焦磷酸盐(IPP)信号分子 1,5-IPP 的细胞内水平敏感。IP 动力学由 Asp1 决定,Asp1 是一种由 N 端 IPP 1-激酶和 C 端 IPP 1-焦磷酸酶结构域组成的双功能酶,分别催化 IP 合成和分解代谢。在这里,我们研究了 Asp1 激酶结构域和全长 Asp1 的活性和特异性。我们发现,Asp1 激酶与 5-IP 的反应速度比与 IP 的反应速度快 22 倍,并且强烈偏向于 IP 合成,而不是在与 IP 反应时看到的显著非生产性 ATP 水解。我们报告说,全长 Asp1 通过其激酶成分催化 1-磷酸的无效磷酸化循环,并通过其焦磷酸酶成分催化 1-焦磷酸水解,导致 ATP 底物的非生产性净消耗。生理浓度的无机磷酸盐选择性拮抗 IP 转化,从而使 1,5-IPP 的净合成成为可能。