CNRS, Université Paul Sabatier, ENFA, UMR 5174 EDB (Laboratoire Évolution & Diversité Biologique), Bât. 4R1, F-31062, Toulouse, France.
Université de Toulouse, UPS, EDB, F-31062, Toulouse, France.
Heredity (Edinb). 2018 Jan;120(1):13-24. doi: 10.1038/s41437-017-0005-6. Epub 2017 Nov 8.
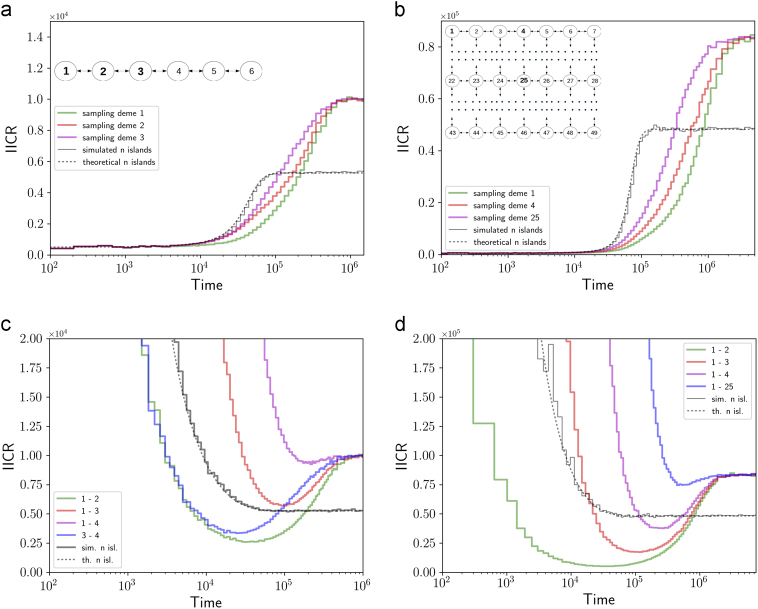
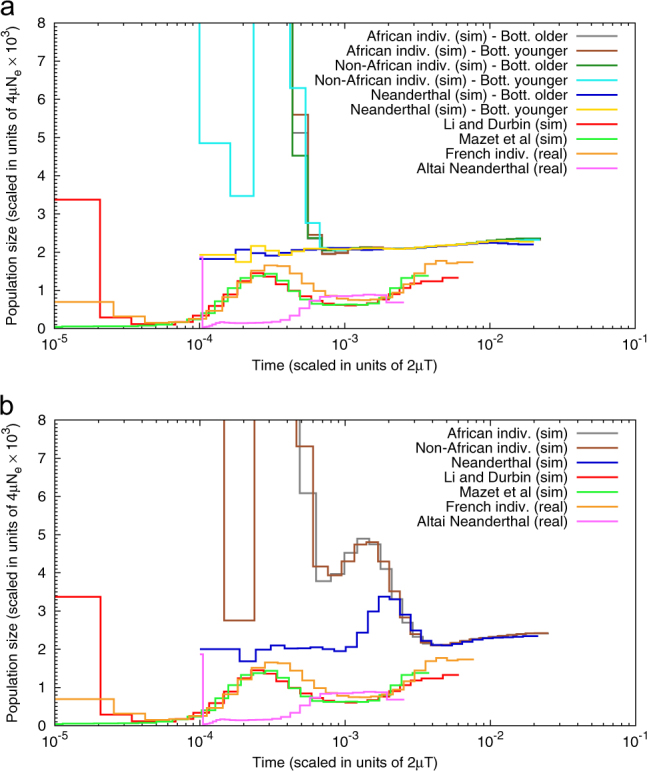
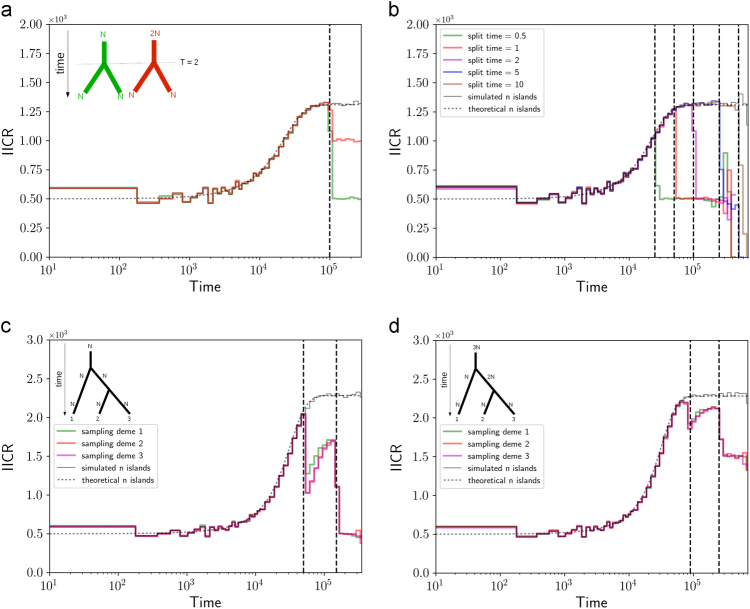
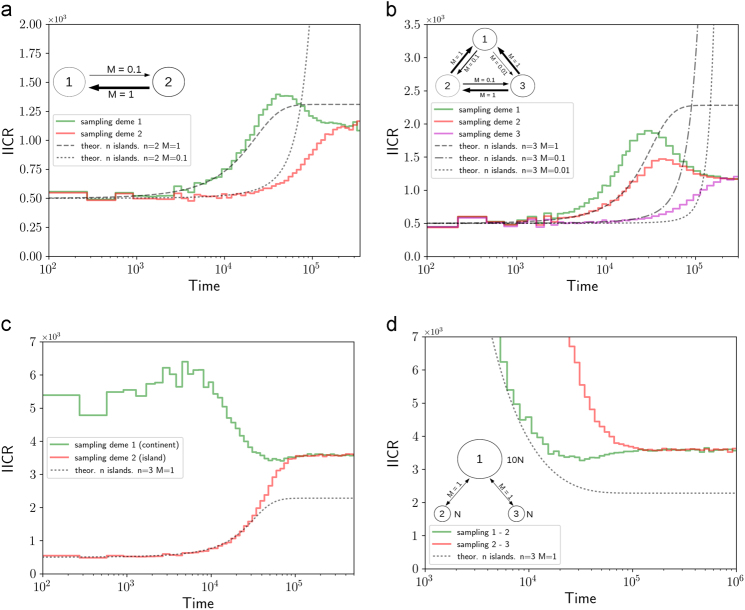
Several inferential methods using genomic data have been proposed to quantify and date population size changes in the history of species. At the same time an increasing number of studies have shown that population structure can generate spurious signals of population size change. Recently, Mazet et al. (2016) introduced, for a sample size of two, a time-dependent parameter, which they called the IICR (inverse instantaneous coalescence rate). The IICR is equivalent to a population size in panmictic models, but not necessarily in structured models. It is characterised by a temporal trajectory that suggests population size changes, as a function of the sampling scheme, even when the total population size was constant. Here, we extend the work of Mazet et al. (2016) by (i) showing how the IICR can be computed for any demographic model of interest, under the coalescent, (ii) applying this approach to models of population structure (1D and 2D stepping stone, split models, two- and three-island asymmetric gene flow, continent-island models), (iii) stressing the importance of the sampling strategy in generating different histories, (iv) arguing that IICR plots can be seen as summaries of genomic information that can thus be used for model choice or model exclusion (v) applying this approach to the question of admixture between humans and Neanderthals. Altogether these results are potentially important given that the widely used PSMC (pairwise sequentially Markovian coalescent) method of Li and Durbin (2011) estimates the IICR of the sample, not necessarily the history of the populations.
已经提出了几种使用基因组数据的推理方法来量化和确定物种历史上的种群大小变化。与此同时,越来越多的研究表明,种群结构会产生种群大小变化的虚假信号。最近,Mazet 等人(2016 年)提出了一种依赖时间的参数,称为 IICR(反向瞬时合并率),用于样本量为二的情况。IICR 在混合模型中等同于种群大小,但在结构模型中不一定如此。它的特点是时间轨迹,表明种群大小随采样方案而变化,即使总种群大小保持不变。在这里,我们扩展了 Mazet 等人(2016 年)的工作,(i)展示了如何在合并的情况下为任何感兴趣的人口统计模型计算 IICR,(ii)将这种方法应用于人口结构模型(1D 和 2D 踏脚石、分裂模型、两岛和三岛不对称基因流、大陆-岛屿模型),(iii)强调采样策略在产生不同历史中的重要性,(iv)认为 IICR 图可以看作是基因组信息的总结,因此可以用于模型选择或排除(v)将这种方法应用于人类和尼安德特人之间混合的问题。总的来说,这些结果非常重要,因为广泛使用的 Li 和 Durbin(2011 年)的 PSMC(成对顺序马尔可夫合并)方法估计的是样本的 IICR,而不一定是种群的历史。