From the Institute of Biophysics, Johannes Kepler University of Linz, Gruberstrasse 40, 4020 Linz, Austria, and.
the Center for Nanobiology and Structural Biology, Institute of Microbiology, Academy of Sciences of the Czech Republic, 373 33 Nove Hrady, Czech Republic.
J Biol Chem. 2018 Jan 26;293(4):1271-1285. doi: 10.1074/jbc.M117.812693. Epub 2017 Dec 13.
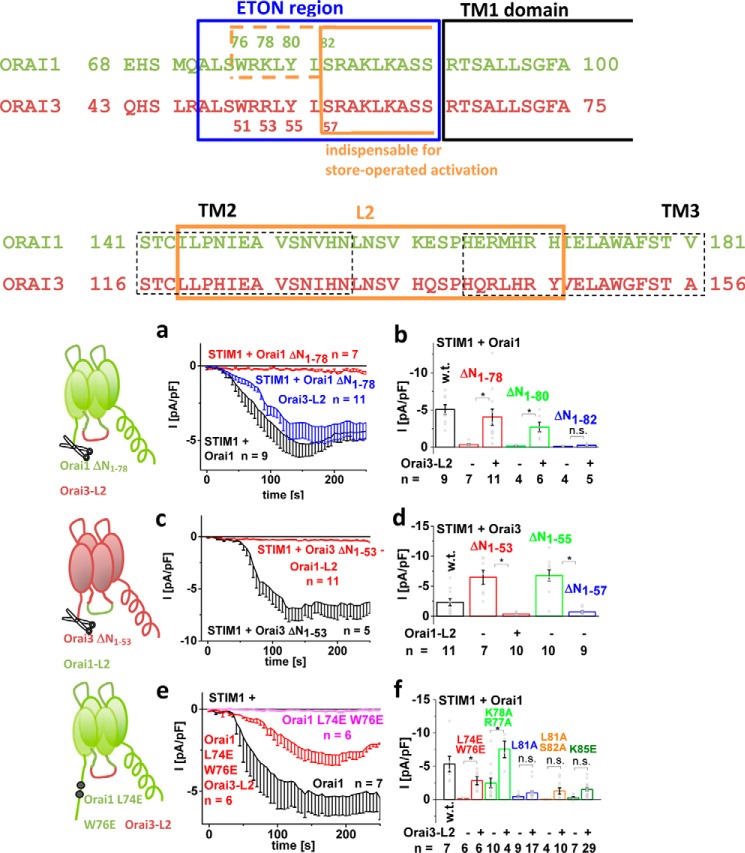
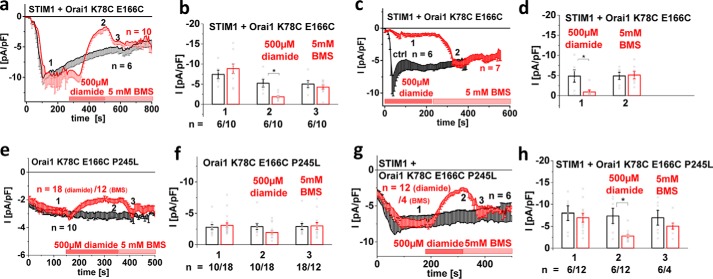
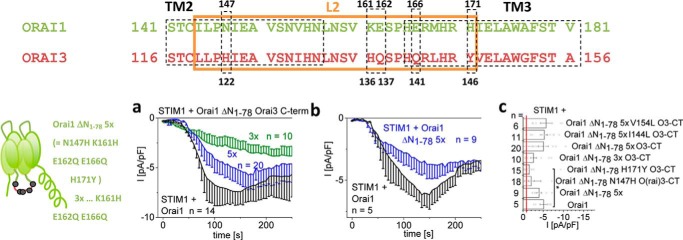
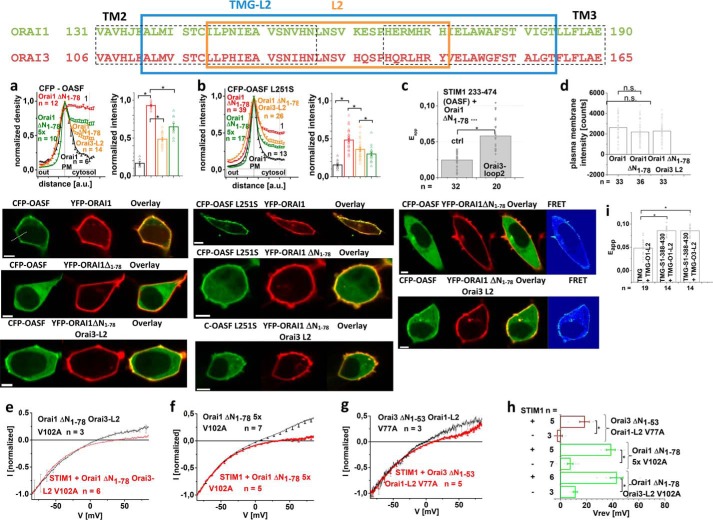
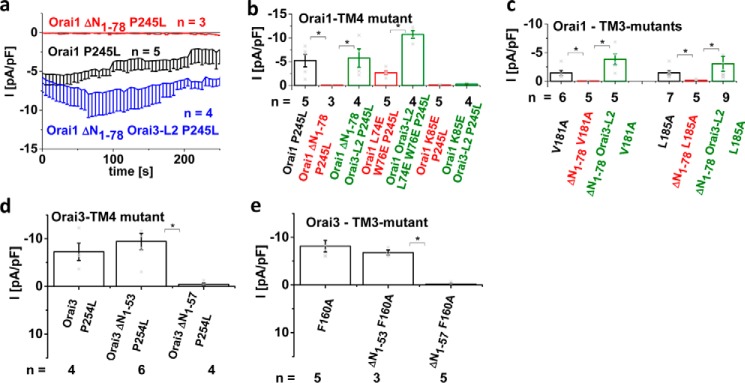
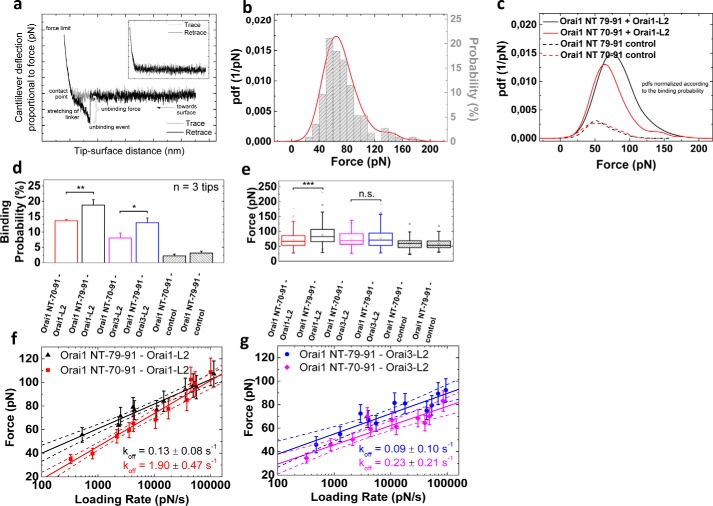
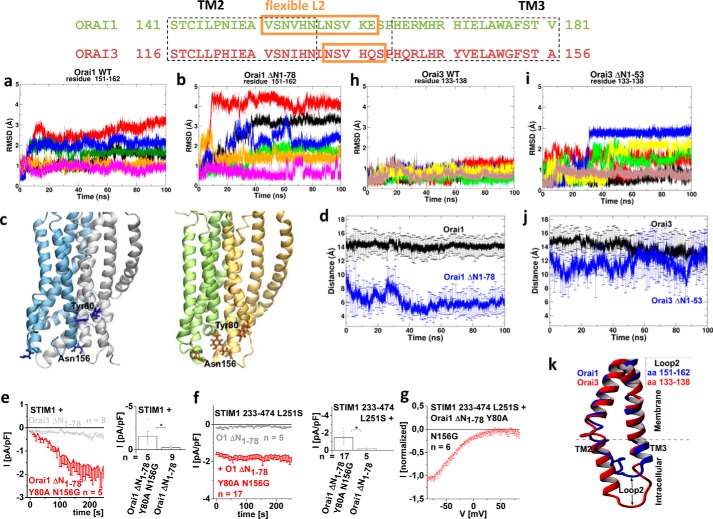
Ca release-activated Ca (CRAC) channels constitute the major Ca entry pathway into the cell. They are fully reconstituted via intermembrane coupling of the Ca-selective Orai channel and the Ca-sensing protein STIM1. In addition to the Orai C terminus, the main coupling site for STIM1, the Orai N terminus is indispensable for Orai channel gating. Although the extended transmembrane Orai N-terminal region (Orai1 amino acids 73-91; Orai3 amino acids 48-65) is fully conserved in the Orai1 and Orai3 isoforms, Orai3 tolerates larger N-terminal truncations than Orai1 in retaining store-operated activation. In an attempt to uncover the reason for these isoform-specific structural requirements, we analyzed a series of Orai mutants and chimeras. We discovered that it was not the N termini, but the loop2 regions connecting TM2 and TM3 of Orai1 and Orai3 that featured distinct properties, which explained the different, isoform-specific behavior of Orai N-truncation mutants. Atomic force microscopy studies and MD simulations suggested that the remaining N-terminal portion in the non-functional Orai1 N-truncation mutants formed new, inhibitory interactions with the Orai1-loop2 regions, but not with Orai3-loop2. Such a loop2 swap restored activation of the N-truncation Orai1 mutants. To mimic interactions between the N terminus and loop2 in full-length Orai1 channels, we induced close proximity of the N terminus and loop2 via cysteine cross-linking, which actually caused significant inhibition of STIM1-mediated Orai currents. In aggregate, maintenance of Orai activation required not only the conserved N-terminal region but also permissive communication of the Orai N terminus and loop2 in an isoform-specific manner.
钙释放激活钙(CRAC)通道构成了细胞内钙离子进入的主要途径。它们通过钙选择性 Orai 通道和钙感应蛋白 STIM1 之间的膜间偶联完全重建。除了 Orai C 末端(STIM1 的主要偶联位点)之外,Orai 通道门控也需要 Orai N 末端。尽管 Orai1 和 Orai3 同工型的跨膜 Orai N 末端延伸区(Orai1 氨基酸 73-91;Orai3 氨基酸 48-65)完全保守,但 Orai3 比 Orai1 耐受更大的 N 末端截短,以保留储存操作激活。为了揭示这些同工型特异性结构要求的原因,我们分析了一系列 Orai 突变体和嵌合体。我们发现,不是 N 末端,而是 Orai1 和 Orai3 的 TM2 和 TM3 之间连接的环 2 区域具有不同的特性,这解释了 Orai N 末端截断突变体的不同、同工型特异性行为。原子力显微镜研究和 MD 模拟表明,无功能的 Orai1 N 末端截断突变体中剩余的 N 末端部分与 Orai1 环 2 区域形成了新的抑制性相互作用,但与 Orai3 环 2 区域没有相互作用。这种环 2 交换恢复了 N 末端截断 Orai1 突变体的激活。为了模拟全长 Orai1 通道中 N 末端和环 2 之间的相互作用,我们通过半胱氨酸交联诱导 N 末端和环 2 之间的接近,这实际上导致了 STIM1 介导的 Orai 电流的显著抑制。总之,Orai 激活的维持不仅需要保守的 N 末端区域,还需要 Orai N 末端和环 2 以特定同工型的方式进行允许的通讯。